LncRNA-ATB promotes autophagy by activating Yes-associated protein and inducing autophagy-related protein 5 expression in hepatocellular carcinoma
2019-10-11ChuanZhuoWangGuangXinYanDeShuoDongHeXinZhaoYuLiu
Chuan-Zhuo Wang, Guang-Xin Yan, De-Shuo Dong, He Xin, Zhao-Yu Liu
Abstract
Key words: LncRNA-ATB; Autophagy; Yes-associated protein; Autophagy-related protein 5; Hepatocellular carcinoma
INTRODUCTION
Hepatocellular carcinoma (HCC) is the second leading cause of cancer-related mortality among males, and is the fifth most common cancer worldwide[1,2]. Although therapy for HCC has seen significant improvement in recent years, clinical outcome prognosis remains poor for patients with HCC. A large number of aberrantly expressed genes influence the progression of HCC, but the molecular mechanisms governing HCC malignancy are still not entirely clear, and the potential connection between long non-coding RNAs (lncRNAs) and autophagy remains to be fully elucidated. There is an important and unmet need to elucidate molecular mechanisms of autophagy, and to capitalize on that knowledge to develop autophagy-related methods as therapeutic strategies for treatment of HCC.
LncRNAs are a class of RNA transcripts that are longer than 200 nucleotides and exhibit limited protein-coding capacity[3]. LncRNAs regulate many aspects of cancer progression and can influence different malignant behaviors, including cancer cell proliferation, apoptosis, metastasis, glycolysis, and angiogenesis[4,5]. LncRNAactivated by transforming growth factor beta (lncRNA-ATB) is a lncRNA transcript regulated by transforming growth factor beta signaling; it mediates induction of epithelial-mesenchymal transition (EMT) downstream of transforming growth factor beta signaling by competitively binding to members of the miR-200 family[6]. Recently,up-regulation of lncRNA-ATB was reported in a variety of human cancers, and was found to influence a multitude of cellular functions in cancer cells[7]. Emerging reports have identified the role of lncRNAs in regulating autophagy[8], but the involvement of lncRNA-ATB in autophagy in HCC is not entirely clear.
Autophagy is an evolutionarily conserved catabolic process that regulates the coordinated lysosomal degradation of cellular components and damaged organelles.Autophagy can support cell survival and maintenance of homeostasis in response to different forms of stress, such as hypoxia, or deprivation of nutrients and energy[9].Additionally, autophagy can promote the invasion and migration of HCC cells[10,11].Nevertheless, whether and how autophagy facilitates cancer progression remains controversial[12]. Based on the cytoprotective properties of autophagy in cancer cells,most research into autophagy has focused on exploring the value of autophagytargeted therapy[13]. There are currently more than 50 randomized controlled trials evaluating the effects of autophagy as relates to cancer therapy. However, the molecular mechanism of autophagy and the exploitation of autophagy as a therapeutic strategy in HCC remain understudied.
In the present study, we evaluated the relationship between lncRNA-ATB and autophagy in HCC. We demonstrated that lncRNA-ATB promotes autophagic flux in HCC cells. We found that lncRNA-ATB regulates autophagy by activating Yesassociated protein (YAP) and increasing autophagy-related protein 5 (ATG5)expression. Our findings provide a novel link between lncRNA-ATB and autophagy,and suggest that lncRNA-ATB may be a potential therapeutic target in the treatment of HCC.
MATERIALS AND METHODS
Patients and tissue samples
Seventy-two HCC tissue samples and adjacent non-tumor tissue samples were obtained from the Cancer Hospital of China Medical University (Shenyang, China).All patients on this study provided informed consent. The human subject research performed in this study was approved by the Clinical Research Ethics Committee of the Hospital of China Medical University. Fresh patient tissue samples were frozen in liquid nitrogen and were immediately stored at -80 °C. The clinical characteristics of the 72 patients with HCC are provided in Table 1.
Cell culture
The human HCC cell lines SMMC-7721 and HepG2 were obtained from the Type Culture Collection of the Chinese Academy of Sciences (Shanghai, China). HCC cells were cultured in RPMI 1640 medium (BioWhittaker, Walkersville, MD, United States)supplemented with 10% fetal bovine serum (FBS; HyClone, Logan, UT, United States)and 1% penicillin/streptomycin in a humidified atmosphere containing 5% CO2at 37°C.
Cell transfection
DNA vectors were transfected into cells using the Lipofectamine 3000 reagent(Invitrogen, CA, United States), according to the manufacturer’s instructions. Briefly,pcDNA3.1 or pcDNA3.1-lncRNA-ATB (Genechem, Shanghai, China) was introduced into cells when cell growth reached approximately 80% confluence. Cells were collected 48 h after transfection. Small interfering RNAs (siRNA) targeting YAP were synthesized by Sigma (Shanghai, China) and the sequences used are as follows: YAP siRNA#1: 5′-GACAUCUUCUGGUCAGAGATT-3′ and YAP siRNA#2: 5′-GGUGAUACUAUCAACCAAATT-3′.
Cell proliferation assay
Proliferation of HCC cells was measured using a Cell Counting Kit-8 (CCK-8,Dojindo, Japan). First, cells transfected with pcDNA3.1-lncRNA-ATB or pcDNA3.1 were seeded at a density of 3000 cells/well into 96-well plates. Cells were cultured at 37 °C for 0, 24, 48, and 72 h. At these indicated time points, 20 μL of CCK8 solution was added to each well. Plates were then incubated at 37 °C for 2 h. Absorbance was measured at 490 nm using a microplate reader. Data are presented as the mean of three independent experiments.
Colony formation assay
For colony formation assays, cells transfected with pcDNA3.1-lncRNA-ATB or pcDNA3.1 were plated in 6-well plates and cultured at 37 °C in growth medium containing 10% FBS. After 14 d, colonies were fixed with 4% polyoxymethylene for 10 min and then stained with 0.1% crystal violet solution for 10 min. Colonies were observed using an Olympus microscope (Tokyo, Japan), and the number of colonies was recorded.
Western blot analysis
Protein lysates were prepared from cells using RIPA buffer supplemented with a protease inhibitor cocktail (Roche, China). Protein lysates were separated by SDSPAGE and then transferred to PVDF membranes (Sigma, United States). Membranes were then incubated with specific antibodies. Protein expression was assessed using ECL chemiluminescent reagents. The primary antibodies used are as follows:antibodies against ATG5, LC3, and β-actin were purchased from Cell Signaling Technology (United States) and those against phospho-PI3K (Tyr458), PI3K, phospho-AKT (Ser473), AKT, phospho-mTOR (Ser2248), mTOR, phospho-YAP (S127), and YAP were purchased from Abcam (United Kingdom).

Table 1 Correlation between lncRNA-ATB expression and clinicopathological parameters in patients with hepatocellular carcinoma
Immunofluorescence staining
For immunofluorescence, cells were fixed with 4% paraformaldehyde (Sigma-Aldrich,United States) for 30 min at room temperature. Cells were then permeabilized with 0.4% Triton X-100 for 5 min at room temperature, and then blocked in 5% bovine serum albumin at 37 ºC for 25 min. Next, cells were incubated with primary antibodies at 4 °C overnight. Following overnight incubation, cells were incubated with specific secondary antibodies for 1 h at 37 °C, and were then washed with PBS three times. After a final wash, nuclei were stained with DAPI for 3 min at room temperature. Immunofluorescence was observed and evaluated using a confocal microscope (Olympus, United States).
Transmission electron microscopy (TEM)
HCC cell samples were processed and autophagosomes were visualized by TEM as described previously[14]. In brief, samples were fixed with 2.5% glutaraldehyde in 0.1 mol/L cacodylate buffer for 4 h and underwent post-fixation with 1% OsO4in 0.1 mol/L cacodylate buffer for 2 h. Next, samples were dehydrated, embedded in resinpropylene oxide, and sectioned using a Leica UFC6 ultra-thin microtome at 80 nm thickness. Finally, images were captured using a Hitachi Model H-7650 transmission electron microscope.
RNA isolation and real time-PCR
Isolation of total RNA, cDNA reverse transcription, and quantitative real-time PCR were performed as previously described[10]. The PCR amplification primers used are as follows: LncRNA-ATB forward, 5’-CTTCACCAGCACCCAGAGA-3’ and reverse,5’-AAGACAGAAAAACAGTTCCGAGTC-3’; GAPDH forward, 5’-AAAGATGTG CTTCGAGATGTGT-3’ and reverse, 5’-CACTTTGTCAGTTACCAACGTCA-3’; ATG3 forward 5’-GACCCCGGTCCTCAAGGAA-3’ and reverse, 5’-TGTAGCCCA TTGCCATGTTGG-3’; ATG5 forward, AAAGATGTGCTTCGAGATGTGT-3’ and
reverse, 5’-CACTTTGTCAGTTACCAACGTCA-3’; ATG7 forward 5’-CAGTTTGCCC CTTTTAGTAGTGC-3’ and reverse, 5’-CCAGCCGATACTCGTTCAGC-3’; ATG10 forward 5’-AGACCATCAAAGGACTGTTCTGA-3’ and reverse, 5’-GGGTAGATGC TCCTAGATGTGAC-3’; ATG12 forward 5’-CTGCTGGCGACACCAAGAAA-3’ and reverse, 5’-CGTGTTCGCTCTACTGCCC-3’; ATG16L forward 5’-AAGAAAC GTGGGGAGTT AGC-3’ and reverse, 5’-AGAGACAGAGCGTCTCCCAA -3’.
RNA immunoprecipitation (RIP)
HepG2 and SMMC-7721 cells were co-transfected with pcDNA3.1-MS2, pcDNA3.1-ATB-MS2, pcDNA3.1-ATB-MS2-mut (ATG5), and pMS2-GFP. The plasmids were synthesized by Genechem (Shanghai, China). After transfection for 48 h, cells were cultured to use in experiments for RIP. RIP was performed using a GFP antibody and the Magna RIPTM RNA-Binding protein, according to the manufacturer’s instructions.
Statistical analysis
All of data analyses were performed using SPSS software (version 17.0, SPSS). Data are presented as the mean ± SD. Significant differences between groups were analyzed using the Student’s t-test. Chi-square tests were performed to determine the relationship between lncRNA-ATB and clinicopathological characteristics. Pearson’s correlation analysis was used to evaluate correlations in expression between two genes. Kaplan-Meier survival analysis was used to evaluate overall survival, and the log-rank test was used to determine differences in survival between groups.Differences were considered to be statistically significant ataP < 0.05 andbP < 0.01.
RESULTS
LncRNA-ATB is highly expressed in HCC and is predictive of poor prognosis
We evaluated expression profiles of lncRNA-ATB in HCC samples by measuring lncRNA-ATB levels in 72 pairs of HCC and adjacent non-tumor tissues by qRT-PCR.LncRNA-ATB was expressed at higher levels in HCC tissues than in non-tumor tissue(Figure 1A). High expression of lncRNA-ATB was associated with larger tumor size and advanced TNM stage. The expression of lncRNA-ATB in patients with HCC tumors larger than 5 cm was significantly higher than in patients with tumor size smaller than 5 cm (Figure 1B). LncRNA-ATB was significantly higher in HCC patients with advanced TNM stage (III/IV) than in patients with local TNM stage (I/II)(Figure 1C). We stratified the 72 patients into a high lncRNA-ATB group and a low lncRNA-ATB group, based on the median expression of lncRNA-ATB. We then examined the relationship between lncRNA-ATB expression and prognosis in patients with HCC. Kaplan-Meier survival analysis revealed that patients with higher lncRNA-ATB expression had a poorer prognosis (Figure 1D). These results indicated that lncRNA-ATB is overexpressed in HCC compared to normal tissues, and that higher lncRNA-ATB expression predicts poor survival for patients with HCC.
Overexpression of lncRNA-ATB promotes HCC cell proliferation and clonogenicity
To explore the function of lncRNA-ATB in HCC cells, we established lncRNA-ATB overexpressing cell lines by transfecting SMMC-7721 and HepG2 cells with pcDNA3.1-ATB. At 48 h post-transfection, lncRNA-ATB expression was about 29-fold and 34-fold higher than that in cells transfected with negative control (pcDNA3.1),respectively (Figure 2A). Overexpression of lncRNA-ATB significantly promoted HCC cell proliferation, as indicated by the CCK-8 proliferation assay (Figure 2B). In vitro colony formation assays demonstrated that overexpression of lncRNA-ATB increased the number of colonies formed by both SMMC-7721 and HepG2 cells(Figure 2C). These results show that lncRNA-ATB has an oncogenic capacity to facilitate the proliferation and clonogenicity of HCC cells.
LncRNA-ATB promotes autophagic flux in HCC cells
To determine the biological effect of lncRNA-ATB on autophagy, we evaluated the effects of lncRNA-ATB overexpression on autophagy in HCC cells. Overexpression of lncRNA-ATB increased the accumulation of LC3-II, which is generally considered to be a marker of autophagosome formation. Furthermore, we used bafilomycin A1, an inhibitor of the fusion of autophagosomes and lysosomes, to examine autophagic flux by evaluating changes in the degradation of LC3-II. Accumulation of LC3-II was significantly increased in HCC cells treated with bafilomycin A1, after transfection with pcDNA3.1-lncRNA-ATB (Figure 3A and B), indicating that lncRNA-ATB induces autophagic flux. By using a LC3-GFP reporter, the abundance of LC3 puncta was measured, and HCC cells transfected with pcDNA3.1-lncRNA-ATB exhibited similar LC3 puncta accumulation to cells treated with rapamycin (Figure 3C and D). TEM analysis detected autophagosomes and autolysosomes in HCC cells overexpressing lncRNA-ATB, and fewer were observed in negative control cells (Figure 3E and F).These results indicate that lncRNA-ATB promotes autophagic flux and autolysosome formation.

Figure 1 LncRNA-ATB is significantly up-regulated in human hepatocellular carcinoma tissues and predicts a poor prognosis. A: Relative expression levels of lncRNA-activated by transforming growth factor beta (lncRNA-ATB) in hepatocellular carcinoma (HCC) tissues and adjacent normal tissues; B: Relative expression levels of lncRNA-ATB in HCC patients with larger tumor sizes (> 5 cm) and smaller tumor sizes (≤ 5 cm); C: Relative expression levels of lncRNA-ATB in HCC patients with early stage (I/II) and advanced stage (III/IV) disease; D: Kaplan-Meier analysis of overall survival in 72 HCC patients according to median lncRNA-ATB expression. The log-rank test was used to calculate the P-value. Data are expressed as the mean ± SD of three independent experiments. Statistical significance is indicated at aP < 0.05. HCC: Hepatocellular carcinoma; LncRNA-ATB: LncRNA-activated by transforming growth factor beta.
LncRNA-ATB promotes autophagy by modulating YAP activation
YAP is a key coactivator of the Hippo pathway, and has been demonstrated to promote cancer cell survival by enhancing autophagic flux[15]. Moreover, a growing number of studies have found that lncRNAs are able to regulate Hippo-YAP signaling in cancer cells. We therefore hypothesized that lncRNA-ATB promotes autophagy by activating YAP. To explore this, we measured YAP expression in HCC cells overexpressing lncRNA-ATB, and found that overexpression of lncRNA-ATB did not change the expression of total YAP protein, but decreased p-YAP expression in SMMC-7721 and HepG2 cells (Figure 4A and B). In addition, consistent with the decrease in p-YAP, overexpression of lncRNA-ATB induced YAP translocation from the cytoplasm to the nucleus, as indicated by immunofluorescence (Figure 4C and D).These data demonstrate that lncRNA-ATB induces nuclear translocation of YAP. To determine whether lncRNA-ATB promotes autophagy by activating YAP, we inhibited YAP expression with siRNA. YAP expression was significantly decreased by si-YAP1, as demonstrated by Western blot analysis (Figure 4E and F). Consistently,we found that YAP knockdown partially attenuated lncRNA-ATB-induced activation of autophagy in HCC cells (Figure 4G and H). These results reveal that lncRNA-ATB promotes autophagy in HCC cells by modulating YAP activation.
LncRNA-ATB influences autophagy by targeting ATG5 expression

Figure 2 Overexpression of lncRNA-ATB promotes the proliferation of hepatocellular carcinoma cells in vitro. A: LncRNA-activated by transforming growth factor beta (lncRNA-ATB) expression was determined by real-time PCR in hepatocellular carcinoma (HCC) cells transfected with pcDNA3.1-lncRNA-ATB; B: CCK-8 assays were performed to determine the proliferation of HCC cell lines (SMMC-7721 and HepG2) transfected with pcDNA3.1-lncRNA-ATB; C: Colony formation assays were performed to measure the proliferation of HCC cells transfected pcDNA3.1-lncRNA-ATB. Data are expressed as the mean ± SD of three independent experiments. Statistical significance is indicated at aP < 0.05 or bP < 0.01. HCC: Hepatocellular carcinoma; LncRNA-ATB: LncRNA-activated by transforming growth factor beta.
Autophagy is a dynamic, continuous, and tightly coordinated metabolic process that is regulated by a series of autophagy-related genes (ATGs). ATG proteins are essential for the formation of the autophagosome, and are crucial for delivery of autophagic cargo to fuse with the lysosome[16]. To determine which ATG genes are involved in regulation of autophagy by lncRNA-ATB, we used real-time PCR to evaluate the expression of ATG genes in HCC cells following overexpression of lncRNA-ATB.ATG5 mRNA expression was significantly up-regulated by lncRNA-ATB overexpression (Figure 5A and B). In addition, Western blot analysis confirmed that ATG5 protein expression was increased by lncRNA-ATB overexpression (Figure 5C and D). Next, we explored the mechanism by which lncRNA-ATB regulates ATG5.LncRNAs can interact with mRNAs to increase mRNA stability, which promotes gene expression[6,17]. We used BLAST to find potential binding sites between lncRNA-ATB and the ATG5 mRNA, and identified several regions of high complementarity. We used RIP to investigate direct interactions of lncRNA-ATB and ATG5 mRNA, and found that LncRNA-ATB was significantly enriched for ATG5 mRNA compared to the empty vector or to lncRNA-ATB with mutated binding sites (Figure 5E and F). We then determined the expression correlation between ATG5 and lncRNA-ATB in HCC tissue. These demonstrate that lncRNA-ATB interacts with ATG5 mRNA and regulates autophagy by increasing ATG5 expression.
Nuclear localization of YAP and ATG5 expression are correlated positively with lncRNA-ATB expression
We examined YAP expression in 72 HCC tissues by immunohistochemistry. We found that 62.5% (45/72) of HCC samples showed stronger YAP staining than adjacent non-tumor tissues (Figure 6A). YAP localization showed stronger nuclear staining in HCC tissues than in normal tissues. We then evaluated ATG5 expression by real-time PCR and found that ATG5 expression was significantly higher in HCC tissues compared with adjacent normal tissues (Figure 6B). Additionally, we determined the correlation between the expression of lncRNA-ATB and ATG5, and found that ATG5 mRNA expression was positively correlated with lncRNA-ATB expression (Figure 6C).

Figure 3 LncRNA-ATB promotes autophagy in hepatocellular carcinoma cells. A and B: Hepatocellular carcinoma (HCC) cells transfected with pcDNA3.1-lncRNA-ATB were subjected to Western blot analysis to detect LC3-II/LC3-I expression, with and without bafilomycin A1 treatment (20 μmol/L for 6 h); C and D:Immunofluorescence of LC3 puncta (green) was measured in HCC cells transfected with pcDNA3.1-lncRNA-ATB or treated with rapamycin; E and F: Electron microscopy identified highly visible autophagosomes and autolysosomes in HCC cells overexpressing lncRNA-ATB compared to negative control cells. HCC:Hepatocellular carcinoma; LncRNA-ATB: LncRNA-activated by transforming growth factor beta.
DISCUSSION
LncRNAs play vital roles in tumor progression[18], and more and more lncRNAs are being shown to influence tumor cells biology[19]. In the present study, we found that lncRNA-ATB is expressed at high levels in HCC tissues. We report that elevated expression of lncRNA-ATB associates with increased tumor size and higher TNM stage, suggesting that lncRNA-ATB may serve as a prognostic biomarker to identify those HCC patients who are at higher risk of disease progression. In previous studies,lncRNA-ATB was found to be overexpressed in various tumors of the digestive system, such as gastric cancer, colorectal cancer, and pancreatic cancer[20-22]. In gastric cancer, Iguchi et al[21]found that expression of lncRNA-ATB significantly correlates with postoperative overall survival of patients with GC, and induced EMT in GC cells. They reported that lncRNA-ATB expression is correlated to tumor size, depth of tumor invasion, and TNM stage in colorectal tumors. In functional experiments,lncRNA-ATB was found to influence multiple biological processes of cancer cells.LncRNA-ATB promotes cell viability, migration, and invasion in T24 cells by regulating miR-126 as a molecular sponge[23]. Here we report that lncRNA-ATB overexpression promotes proliferation of HCC cells. These data suggest that lncRNAATB plays an oncogenic role in HCC progression.
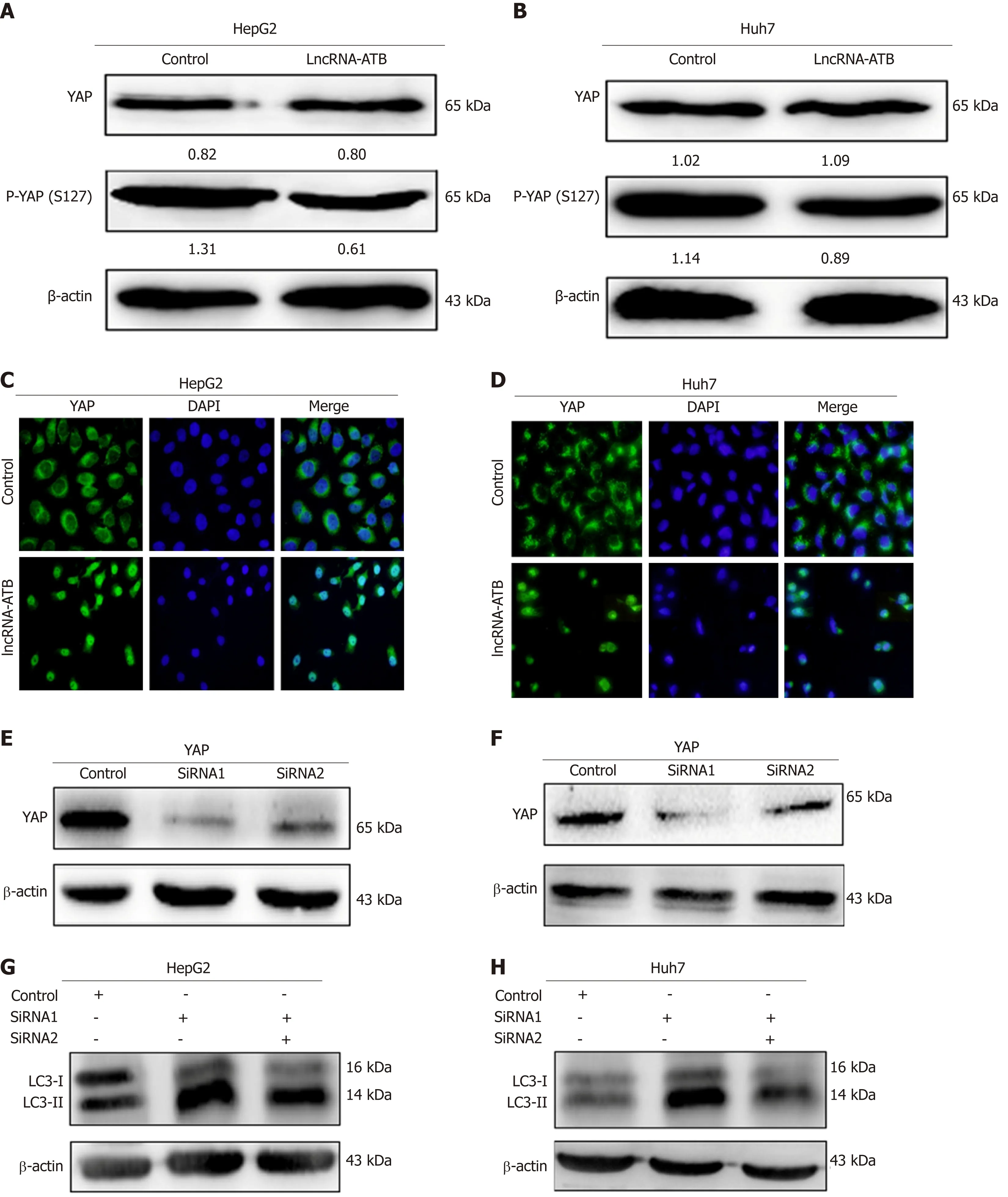
Figure 4 LncRNA-ATB promotes autophagy through modulation of Yes-associated protein activation. A and B: The protein levels of total Yes-associated protein (YAP) and p-YAP in hepatocellular carcinoma (HCC) cells transfected with pcDNA3.1-lncRNA-ATB were measured by Western blot; C and D: The nuclear localization of YAP was detected in transfected HCC cells by immunofluorescence; E and F: Knockdown of YAP in HCC cells transfected with si-YAP (si-YAP 1# and si-YAP 2#) was measured using Western blot; G and H: HCC cells were co-transfected with pcDNA3.1-lncRNA-ATB and si-YAP, and the protein levels of LC3 were determined by Western blot. Yes-associated protein: YAP; HCC: Hepatocellular carcinoma; LncRNA-ATB: LncRNA-activated by transforming growth factor beta.
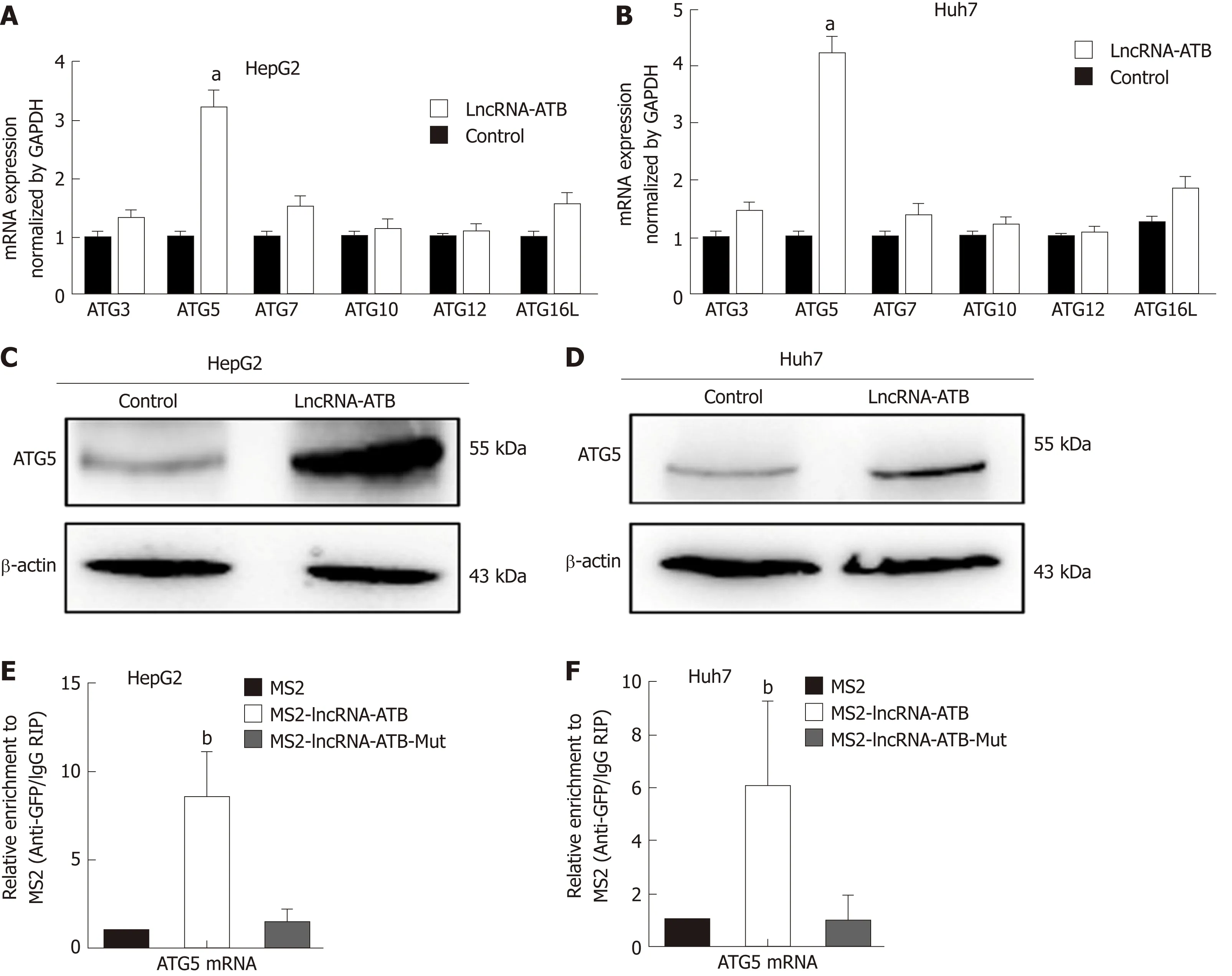
Figure 5 LncRNA-ATB induces autophagy by regulating autophagy-related protein 5 expression. A and B: The mRNA expression of autophagy-related genes involved in autophagy was examined by real-time PCR in hepatocellular carcinoma (HCC) cells transfected with lncRNA-ATB; C and D: The protein levels of ATG5 were analyzed in HCC cells transfected with lncRNA-ATB by Western blot; E and F: RIP-derived RNA was evaluated by real-time PCR. Data are expressed as the mean ± SD of three independent experiments. Statistical significance is indicated at aP < 0.05 or bP < 0.01. ATG3: Autophagy-related protein 3; ATG5: Autophagyrelated protein 5; ATG7: Autophagy-related protein 7; ATG10: Autophagy-related protein 10; ATG12: Autophagy-related protein 12; ATG16L: Autophagy-related protein 16L; HCC: Hepatocellular carcinoma; RIP: RNA immunoprecipitation; LncRNA-ATB: LncRNA-activated by transforming growth factor beta.
Autophagy can be considered a kind of metabolic reprogramming pathway, and is a topic of increasing relevance to cancer research. The role of autophagy as an antitumor or tumor-promoting pathway is context dependent[24], and emerging studies have indicated that autophagy can facilitate survival of tumor cells in response to stress[9,25]. Reports have also demonstrated that non-coding RNAs can act as regulators in the autophagy pathway[8]. In our current study, we analyzed the potential link between lncRNA-ATB and autophagy. We identified enrichment of autophagy signatures in the gene expression data from SMMC-7721 cells engineered to overexpress lncRNA-ATB. We investigated the influence of lncRNA-ATB on autophagy in HCC cells, and found that overexpression of lncRNA-ATB significantly promoted autophagy flux in HCC cells. The Hippo-YAP pathway regulates autophagy and promotes cancer cell survival during nutrient deprivation[15].Moreover, lncRNAs play a vital role in activating YAP signaling[26,27]. Therefore, we explored whether lncRNA-ATB induces autophagy by regulating YAP. We found that lncRNA-ATB overexpression decreased p-YAP expression and induced the nuclear translocation of YAP. Rescue experiments showed that lncRNA-ATB promotes autophagy by modulating YAP activation. Our previous study found Hippo-YAP signaling plays an essential role in liver cancer development. Our present data suggest that YAP activation acts as a mediator between lncRNA-ATB expression and autophagy.
Autophagy is a dynamic and continuous process that is regulated by a series of ATG proteins. Yoshinori Ohsumi’s laboratory identified the core ATGs that are essential for formation of the autophagosome and delivery of autophagic cargo to the lysosome[28]. In addition, lncRNAs have been shown to coordinate autophagy by influencing the expression and post-transcriptional regulation of ATG genes[8]. For example, lncRNA PVT1 activates autophagy in glioma vascular endothelial cells by upregulating the expression of ATG7 and beclin1[29]. In our study, we found that lncRNA-ATB increases ATG5 expression by interacting with ATG5 mRNA. ATG5 is part of the ATG12-ATG5-ATG16L1 complex, and it enhances ATG3-mediated conjugation of ATG8 family proteins, resulting in phagophore elongation[30]. Previous studies have also found that lncRNA HNF1A-AS1 functions as an oncogene and promotes autophagy by regulating ATG5. Our results indicate that lncRNA-ATB promotes autophagy by regulating ATG5 expression.

Figure 6 Yes-associated protein nuclear localization and autophagy-related protein 5 expression are positively correlated with lncRNA-ATB expression. A:Representative immunostaining for Yes-associated protein (YAP) in hepatocellular carcinoma (HCC) and adjacent normal tissues (magnification: ×200 or ×400); B:The expression levels of autophagy-related protein 5 (ATG5) in 72 pairs of HCC tumor and adjacent normal tissues were evaluated by real-time PCR; C: Pearson’s correlation analysis of the relationship between expression of lncRNA-ATB and ATG5 (r = 0.527, P < 0.001). Statistical significance is indicated at bP < 0.01. YAP:Yes-associated protein; ATG5: Autophagy-related protein 5; HCC: Hepatocellular carcinoma; LncRNA-ATB: LncRNA-activated by transforming growth factor beta.
In summary, our study elucidates the role of lncRNA-ATB in regulating autophagy in HCC. We report that lncRNA-ATB is often overexpressed in HCC tissues and acts as an oncogene to facilitate the progression of HCC. Our data show that lncRNA-ATB promotes autophagy by modulating YAP activation. Additionally, we report that lncRNA-ATB interacts with ATG5 mRNA and influences autophagy by increasing the expression of ATG5. Our study provides novel insights into the molecular mechanisms by which lncRNA-ATB regulates autophagy in HCC cells.
ARTICLE HIGHLIGHTSA large number of aberrantly expressed genes influence the progression of HCC, but the molecular mechanisms governing HCC malignancy are still not entirely clear, and the potential connection between lncRNAs and autophagy remains to be fully elucidated. There is an important and unmet need to elucidate the molecular mechanisms of autophagy, and to capitalize on that knowledge to develop autophagy-related methods as therapeutic strategies for treatment of HCC.
Research objectives
To explore the influence of lncRNA-ATB in regulating autophagy in HCC cells and the underlying mechanism.
Research methods
We compared the expression of lncRNA-ATB in 72 HCC tissues and adjacent non-tumor tissues by real-time PCR. The role of lncRNA-ATB in cell proliferation and colony formation was evaluated in vitro. The effect of lncRNA-ATB on autophagy was determined using a LC3-GFP reporter and transmission electron microscopy. Furthermore, the mechanism by which lncRNAATB regulates autophagy was explored by immunofluorescence staining, RNA immunoprecipitation, and Western blot.
Research results
We found that lncRNA-ATB was up-regulated in HCC and lncRNA-ATB expression positively correlated with tumor size, TNM stage, and poor survival of HCC patients. Overexpression of lncRNA-ATB promoted cell proliferation and colony formation in vitro. LncRNA-ATB promoted autophagy by activating Yes-associated protein (YAP). Moreover, lncRNA-ATB interacted with autophagy-related protein 5 (ATG5) mRNA and increased ATG5 expression.
Research conclusions
Our study reveals that lncRNA-ATB regulates autophagy by activating YAP and increasing ATG5 expression. These results provide new insights into the role of lncRNA-ATB in autophagy in HCC.
Research perspectives
Our findings provide a novel link between lncRNA-ATB and autophagy, and suggest that lncRNA-ATB may be a potential therapeutic target in the treatment of HCC.
杂志排行

World Journal of Gastroenterology的其它文章
- How does Helicobacter pylori cause gastric cancer through connexins: An opinion review
- Colorectal cancer: Parametric evaluation of morphological,functional and molecular tomographic imaging
- Sarcopenia and cognitive impairment in liver cirrhosis: A viewpoint on the clinical impact of minimal hepatic encephalopathy
- Significance of tumor-infiltrating immunocytes for predicting prognosis of hepatitis B virus-related hepatocellular carcinoma
- Long non-coding RNA highly up-regulated in liver cancer promotes exosome secretion
- Circular RNA PIP5K1A promotes colon cancer development through inhibiting miR-1273a