不同生态条件对小米黄色素含量的影响
2019-10-10杨延兵陈二影王润丰秦岭尹秀波张会迪黎飞飞管延安
杨延兵,陈二影,王润丰,秦岭,尹秀波,张会迪,黎飞飞,管延安
不同生态条件对小米黄色素含量的影响
杨延兵1,陈二影1,王润丰1,秦岭1,尹秀波2,张会迪1,黎飞飞1,管延安1
(1山东省农业科学院作物研究所/山东省特色作物工程实验室,济南 250100;2山东省农业技术推广总站,济南 250100)
【】谷子籽粒脱壳后为小米,小米黄色素含量是反映小米商品性的重要指标,对于小米的商品品质和营养品质都有重要的影响。解析不同生态条件下小米黄色素含量变化规律,为优质特色谷子品种选育及生产利用提供科学依据。选用华北夏谷区选育的8个优质、特色谷子新品种,于2016—2017在不同生态条件下的5个试点种植,成熟收获后,测定小米的黄色素含量,采用多因素方差分析差异显著性,并对黄色素含量和生育期气温、降水量、日照时数等因素进行相关性分析。年份、品种、地点×年份、品种×年份、地点×品种×年份对黄色素含量影响极显著(<0.01),地点、地点×品种对黄色素含量影响显著(<0.05);其中,品种、地点、品种×地点互作对黄色素含量变异贡献率较大,分别为57.12%、27.57%和6.12%。黄色素平均含量2017年显著高于2016年。济南、德州、济宁试点小米黄色素含量2年的结果均显著高于泰安和临沂黄色素含量。5个试点8个谷子品种2年的黄色素平均含量为23.42 mg·kg-1,变幅为18.56—26.14 mg·kg-1,中谷2号最高,济绿谷1号最低;中谷2号、济糯谷2号、济谷21、济谷19黄色素含量差异不显著,但显著高于豫谷18、济谷22、济谷20、济绿谷1号的黄色素含量。黄色素含量和6月苗期气温、9月灌浆中后期平均气温、生育期平均气温极显著正相关(=0.908,<0.01;=0.798,<0.01;=0.808,<0.01);和9月灌浆中后期的日照时数、生育期日照总时数极显著正相关(=0.771,<0.01;=0.769,<0.01)。不同年份、地点、品种及因素互作对谷子黄色素含量有显著影响,其中,品种因素影响最大;谷子生育期平均气温、降雨量、光照时数等天气因素的变化及时空分布的差异是小米黄色素含量变化的重要原因;谷子灌浆中后期平均气温较高、光照充足有利于黄色素的积累。品种基因型是决定小米黄色素含量的最重要因素。
小米;黄色素含量;生态条件;气温;降雨量;日照时数
0 引言
【研究意义】谷子起源于中国,有10 000年的栽培历史[1-3],被誉为中华民族的哺育作物,对于中华民族的文明延续和发展起到巨大的作用,目前仍是中国北方旱作农业的重要作物。谷子籽粒去壳后为小米,其颜色有黄色、白色、青灰色、黄绿色等,黄色小米品种占谷子品种的90%以上。小米黄色素的主要组分含有叶黄素、玉米黄质以及少量的隐黄质和β-胡萝卜素等类胡萝卜素[4-5],这些类胡萝卜素为天然的抗氧化物质,具有重要的医疗保健价值[6-7],其含量的高低不仅与其营养价值有关,而且是反映小米商品品质的重要指标[8],成为经销商和消费者选择小米的首要指标。研究不同生态条件下小米黄色素含量的变化,解析提高小米产品黄色素含量的途径,对于优质品种选育和区域化布局,发挥品种的提质增效潜力具有重要的意义。【前人研究进展】黄色素含量高低是小麦、玉米、高粱、谷子等禾本科作物非常重要的品质性状,有关黄色素及组分类胡萝卜素相关研究较多[9-18],尤其在小麦、玉米上。这些研究多侧重于对作物资源进行鉴定评价、基因定位、优异基因挖掘等方面。谷子研究上,Yang等[19]等研究了来自华北夏谷区、东北春谷区、西北早熟春谷区、西北中晚熟春谷区等不同生态区的270份品种,发现来自不同生态区的谷子品种黄色素含量具有一定的差异,来自西北中晚熟春谷区的品种小米黄色素含量较高,西北早熟区品种小米黄色素含量较低,小米黄色素含量和生育期显著正相关。Liu等[20]分析了中国育成品种的类胡萝卜素含量;董倩楠等[21]研究了不同施肥条件对谷子β-胡萝卜素和叶黄素的影响;Yano等[22]研究表明,谷子籽粒的黄色素主要在胚乳中积累,谷糠和谷壳中很少,成熟期籽粒中黄色素水平逐渐升高,过度成熟后迅速下降。Ning等[23]研究认为喷施外源硒能显著提高谷子的黄色素含量。这些研究主要集中在对谷子品种资源黄色素的鉴定评价,肥料以及微量元素处理对黄色素含量及组分的影响等方面。【本研究切入点】将小米黄色素含量作为一种品质指标,探讨生态环境因素和小米黄色素含量变化的关系,以及不同生态条件下小米黄色素含量变化规律的研究尚缺乏。【拟解决的关键问题】本研究利用华北夏谷区育成的几个优质、特色米品种,在山东省不同生态条件下研究年份、地点、品种等因素对小米黄色素含量的影响,并对小米黄色素含量和谷子生育期光、温、水等生态因子进行相关性分析,解析小米黄色素含量变化规律及提高小米黄色素含量的途径,为优质谷子品种的选育及生产利用提供科学依据。
1 材料与方法
1.1 试验设计
选取当前山东谷子生产中重点应用种植的8个谷子品种为供试材料(表1),于2016—2017年在山东省不同生态条件下的谷子主要产区进行试验,试验点包括济南(山东农业科学院作物研究所试验基地,E117.09º/N36.71º,海拔24 m)、济宁(山东圣丰种业试验地,E115.01º/N35.42º,海拔56 m)、临沂(临沂市农业科学研究院试验地,E118.26º/N35.09º,海拔51 m)、泰安(泰安市农业科学研究院试验基地,E116.46º/N36.14º,海拔85 m)、德州(德州市农业科学研究院试验地,E116.33º/N37.34º,海拔22 m),各试验点2016、2017谷子生育期6—9月的主要气象要素(平均气温、降雨量、光照时数)来源于山东气象服务网(http://112.33.4.68:9090/index.php)(表2)。
每个品种种植6行,行长5 m,行距0.5 m,留苗密度4.0万/666.7 m2,3次重复,随机区组排列,调查抽穗期、成熟期等物侯期性状。管理措施高于普通大田。成熟时试验小区全收获记产,收获时每小区取样20株,调查主要农艺性状,籽粒脱壳后测定小米黄色素含量。各试验点于当年的6月中旬播种,9月下旬收获,具体时间见表3。
1.2 谷子籽粒小米黄色素含量测定
参照杨延兵等[24]方法测定谷子籽粒黄色素含量。
1.3 数据处理与分析
用Microsoft Excel 2010处理数据,DPS v7.05数据处理系统进行方差分析和差异显著性检验(Duncan新复极差法),利用SPSS22.0进行相关性分析(Pearson法)。
2 结果
2.1 黄色素含量的多因素方差分析和因素贡献率
不同年份、种植地点、品种以及各因素的互作都对小米黄色素含量有显著或极显著影响(<0.05或<0.01)。分析各因素对小米黄色素含量的贡献率发现,品种因素变异贡献率最大,占57.12%,表明品种本身的遗传因素对黄色素含量变异影响最大,其次是试验地点贡献率27.57%,再次是品种×地点互作占6.12%,而不同年份之间及其他因素互作贡献率相对较小(表4)。因此,提高小米黄色素含量应当重视品种和种植区域选择,以便发挥品种的最大潜力。

表1 试验材料名称、类型及选育单位
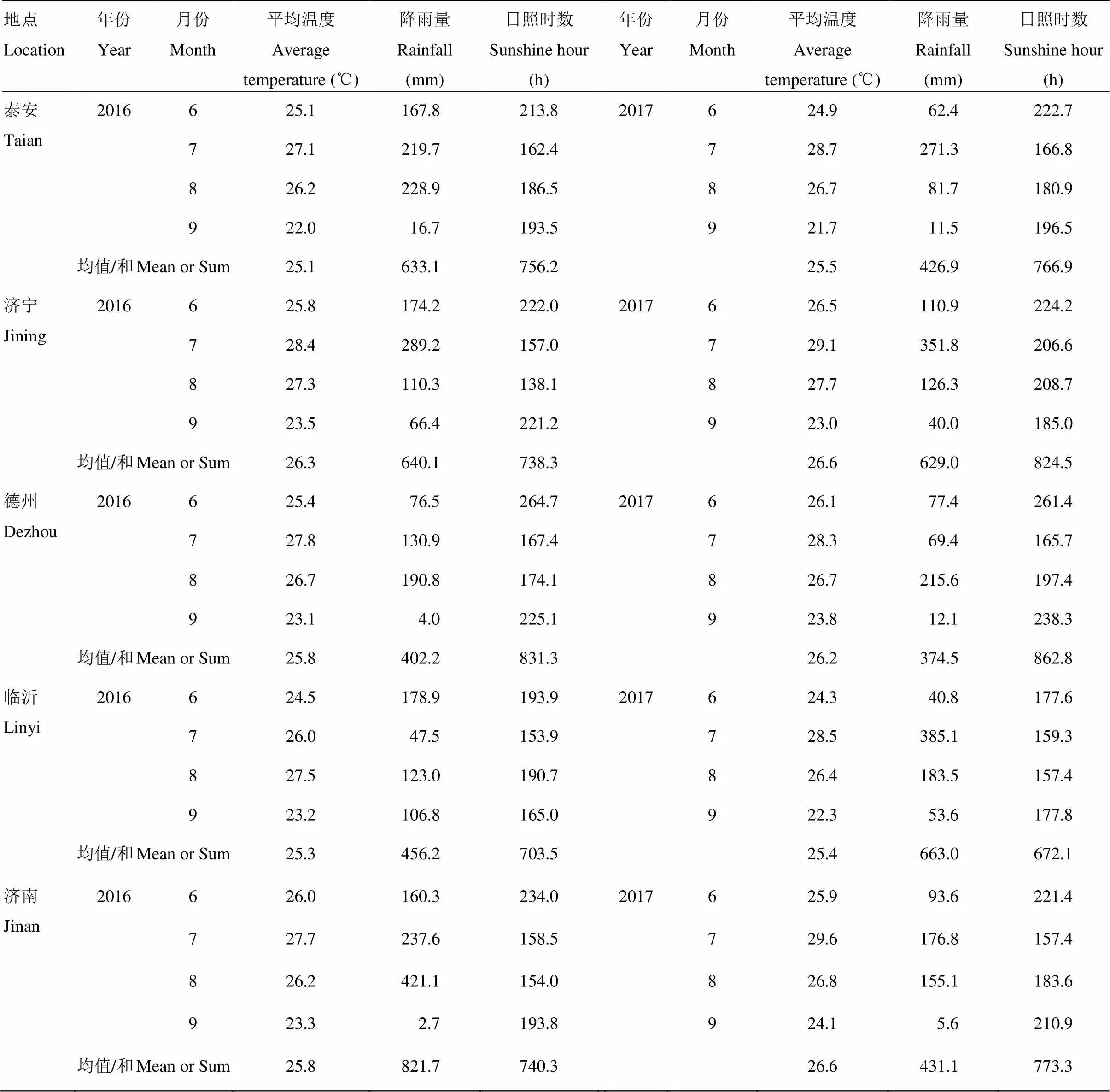
表2 不同试验地点的温度、降雨、日照时数

表3 不同试验地点播种、收获时间和生育总天数

表4 小米黄色素含量的多因素方差分析
贡献率(%)=变因×100%/(总-误-区组) Contribution rate (%)=F×100%/(T-E-B)
2.2 年份、试验地点对小米黄色素含量的影响
不同年份、地点黄色素含量如表5。2016—2017年2年5个试点8个品种黄色素含量平均为23.42 mg·kg-1,2017年黄色素平均含量比2016年平均含量高0.89 mg·kg-1。德州、济南试点年份间黄色素含量差异较大,2017年比2016年分别高15.5%和3.38%;济宁、泰安试点小米黄色素含量2017年比2016年分别高1.56%和1.24%;而临沂试点年份之间差异较小,2017年比2016年减少0.35%。
试验地点对黄色素含量影响显著,济南、德州、济宁试验点2年的黄色素含量水平均显著高于泰安、临沂试验点黄色素含量(<0.01)。济南、济宁、德州试点对于黄色素含量具有正向效应,这三个试验点黄色素含量比平均值分别高1.19、1.06和1.91 mg·kg-1,提高幅度5.08%、4.52%和8.15%;泰安、临沂试点具有负向效应,分别比平均值低1.61和2.54 mg·kg-1,降低幅度6.87%和10.85%。德州、济南、济宁试点2年平均黄色素含量差异不显著,泰安、临沂试点黄色素含量差异不显著。各试点黄色素含量2年间表现不一致,2016年济南、济宁试点显著高于其他试点,2017年德州试点显著高于其他试点。综合2年试验结果,德州试验点黄色素含量最高,临沂试点黄色素含量最低。
2.3 品种之间小米黄色素含量的差异
不同品种小米黄色素含量及变异系数如表6。2年5个试点8个品种黄色素平均含量23.42 mg·kg-1,变幅为18.56—26.14 mg·kg-1,济绿谷1号黄色素含量最低,中谷2号黄色素含量最高。中谷2号、济糯谷2号、济谷21、济谷19黄色素含量显著高于豫谷18、济谷22、济谷20、济绿谷1号(<0.05),而中谷2号、济糯谷2号、济谷21、济谷19黄色素含量品种之间差异不显著,豫谷18和济谷22黄色素含量差异不显著。不同品种黄色素含量高低2年表现趋势基本一致,总体都表现为中谷2号>济糯谷2号>济谷21>济谷19>豫谷18>济谷22>济谷20>济绿谷1号,说明不同品种的黄色素含量变化趋势年份之间相对稳定,品种本身遗传因素影响较大。

表5 不同年份、地点小米黄色素含量的差异
表中数据为平均值;同列数据后不同小写字母表示处理间差异达5%显著水平。下同
Means of values followed by different small letters in the same column mean significant difference at 0.05 level. The same as below
8个品种黄色素含量变异度为1.20%—5.79%,济谷21变异度最大,其次为中谷2号、济谷19;济谷22变异度最小。中谷2号、济糯谷2号、济谷21、济谷19其黄色素变异度略高于其他4个品种。黄色素含量相对较高的品种变异度相对较大。
2.4 品种、地点、年份因素互作对小米黄色素含量的影响
8个品种在5个试点2年的黄色素含量及差异显著性如表7。品种在各地点、年份间表现不一致,黄色素含量的高低顺序不同。中谷2号在泰安、济宁试点2016年、2017年黄色素含量都是最高的,中谷2号在济宁试点2年黄色素含量差2.63 mg·kg-1,差异较大,而其他品种则相对较小。德州试点各品种黄色素含量2017年都高于2016年,中谷2号和豫谷18分别相差0.27和0.77 mg·kg-1,差异相对较小;其他济谷19、济谷20、济谷21、济糯谷2号、济绿谷1号、济谷22相差分别为3.13、4.18、4.16、4.67、3.40和3.96 mg·kg-1,黄色素含量差异较大;济谷21在德州试点2017年小米黄色素含量最高达到31.07 mg·kg-1。临沂试点2016年济糯谷2号黄色素含量最高,而2017年则是中谷2号最高。济谷19、济谷21、济糯谷2号在济南试点连续2年黄色素含量都是最高的,且差异不显著;这三个品种黄色素含量显著高于其他5个品种;济谷19、济谷21、济糯谷2号都是山东省农业科学院作物研究所选育,在本地试验点含量最高,说明品种对本地条件的适应性。济绿谷1号在所有试点黄色素含量在所有品种都是最低的。总之,每个品种都有一定的区域适应性,因此选择适合的地点才能发挥最大的潜力。
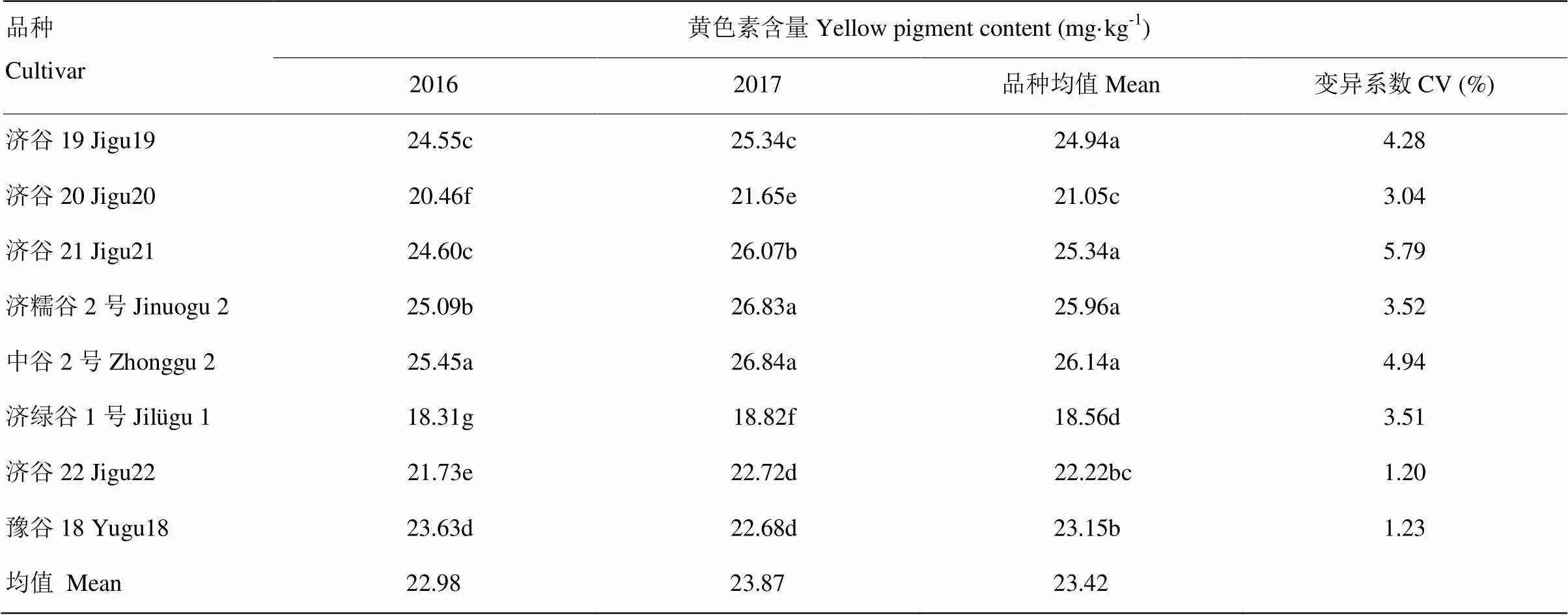
表6 不同品种小米黄色素含量的差异及变异度
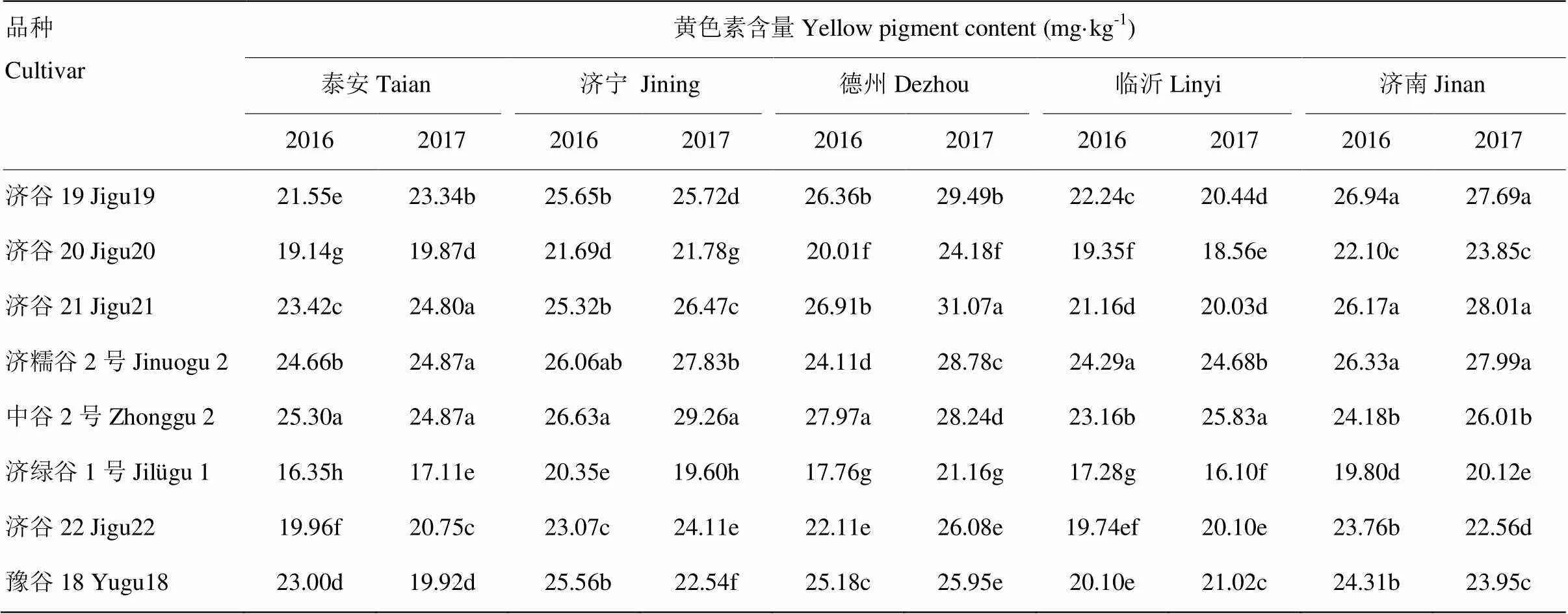
表7 不同品种5个试点2016、2017年的小米黄色素含量的差异
2.5 小米黄色素含量和气象因子相关性
对黄色素含量和各气象因子进行相关性分析(表8),黄色素含量和6月份平均气温、9月份平均气温、生育期平均气温呈极显著正相关(<0.01);与6月、7月、9月降水量及总降水量呈不显著负相关;与6月份光照时数显著正相关(<0.05),与9月份光照时数极显著正相关(<0.01),与生育期光照总时数极显著正相关(<0.01)。

表8 小米黄色素含量和平均气温、降水量、日照时数相关性
*和**表示处理间差异达5%和1%显著水平;YPC:黄色素含量
* and ** represent significances at 0.05 and 0.01 levels; YPC: Yellow pigment content
3 讨论
3.1 品种因素对小米黄色素含量的影响
本研究表明品种之间黄色素含量差异显著,8个品种黄色素含量平均23.42 mg·kg-1,变幅为18.56—26.14 mg·kg-1。据Yang等[13]用270份谷子材料研究,黄色素含量变幅为1.91—28.54 mg·kg-1,平均含量17.80 mg·kg-1,略低于本研究的黄色素含量。本研究黄色素含量平均水平相对较高,主要与研究选用的都是优良品种有关。其中有5个品种在全国优质米鉴评中荣获“一级优质米”,1个品种二级优质米,2份材料为特殊米类型;济糯谷2号为糯质类型,品种本身黄色素含量相对较高;济绿谷1号米色黄绿色,黄色素含量略低。本研究中,中谷2号、济糯谷2号、济谷21、济谷19黄色素含量差异不显著,但显著高于济谷20、济谷22、豫谷18、济绿谷1号。品种对黄色素含量变异贡献率为57.12%,因此,尽管不同年份、不同地点之间存在差异,品种本身的遗传因素才是决定因素。要获得黄色素含量较好的小米产品,品种是首要因素,在品种选育和谷子生产上,应当进一步挖掘优异的基因资源,加强小米黄色素代谢途径有关基因研究,加快培育更多黄色素含量较高的优异品种应用于生产。目前,谷子基因组测序已经完成[25-26],Jia等[27]建立了谷子的单倍型物理图谱;利用全基因组学关联分析也初步定位了和黄色素含量相关的主效基因八氢番茄红素合成酶PSY[28],并开发了与黄色素含量相关的分子标记,通过生物技术改良谷子,提高谷子的黄色素含量成为可能。
3.2 不同年份气象因子的变化对小米黄色素含量的影响
本研究表明不同年份之间小米黄色素含量差异显著,除临沂外,其他试点都表现为2017年黄色素含量高于2016年黄色素含量。其中,德州试点年份之间黄色素含量差异较大,2017年比2016年高3.07 mg·kg-1,其他试点年份之间差异相对较小。黄色素含量和气象因素相关性分析表明,黄色素含量和生育期平均气温极显著正相关(=0.808,<0.01),与生育期光照时数极显著正相关(=0.769,<0.01),尤其与9月份谷子灌浆中后期气温、光照时数极显著正相关(=0.798,<0.01;=0.771,<0.01)。2017年德州试点谷子生育期平均气温、降雨、日照分别为26.2℃、374.5 mm、862.8 h,2016年分别为25.8℃、402.2 mm、831.3 h,2017年比2016年平均气温高0.4℃,日照时数长31.5 h,9月份日照时数长13.2 h,降雨量2016年比2017年多27.7 mm。济南、泰安的情况相似,2017年黄色素含量略高于2016年黄色素含量,其天气条件都表现为平均气温、日照时数2017年高于2016年,而降雨量2017年远低于2016年;济南试点平均气温2017年比2016高0.8℃,日照时数多33 h,降雨少242.0 mm;泰安试点谷子生育期间平均气温2017年比2016年高0.4℃,日照时数长73.9 h,而降雨少290.6 mm。济宁试点平均气温2017年和2016年高0.3℃,降雨相差25.9 mm,光照时数多2017年比2016年多86.2 h。而临沂试点2017年黄色素含量比2016年略低0.06 mg·kg-1,差异较小;临沂试点生育期降雨量2017年比2016年高206.8 mm,而光照2017年比2016年少31.4 h,较多的降雨、阴雨天较多,日照时数短可能是该试点2017年黄色素含量略低于2016年的主要原因。
综上,年份之间谷子生育期平均气温、降雨量、光照时数是影响谷子黄色素含量的重要因素,较高的平均气温、较长的光照、相对较少的降雨量有利于种子中黄色素积累。在小米生产上,应尽可能选择谷子灌浆期光照充足、降雨较少、气温较高的生态环境种植谷子,生产出更加优质的小米。
较高的气温、较长的光照,种子中会积累较多的黄色素,其原因可能是植物的种子中类胡萝卜素是抗氧化系统和捕光色素的组成部分,具有限制自由基对质膜的损伤和抗衰老的功能[29-30],较高的类胡萝卜素水平可能是种子对较高气温和较长光照一种响应。
3.3 地点对小米黄色素含量的影响
地点的改变,不仅涉及到光、温、水等气象因素的变化,还包括不同地点间土壤质地、肥力水平以及耕作栽培制度等多方面的因素,不同地点间小米黄色素含量的变化是综合因素作用的结果。本研究中,济南、德州、济宁试点的黄色素含量显著高于泰安试点和临沂试点黄色素含量。相关性分析表明平均气温的高低和黄色素含量显著正相关,较高的平均气温有利于黄色素的积累。2016、2017年济南、德州、济宁三地谷子生育期平均气温分别为25.8℃(济南)、25.8℃(德州)、26.3℃(济宁)和26.6℃(济南)、26.2℃(德州)、26.6℃(济宁),明显高于泰安、临沂2016、2017年平均气温25.1℃(泰安)、25.3℃(临沂)和25.5℃(泰安)、25.4℃(临沂)。另一方面小米黄色素含量的高低和9月谷子灌浆中后期的光照极显著正相关,2016年9月份济南、德州、济宁光照分别为193.8、225.1和221.2 h明显高于泰安和临沂的193.5和165 h,2017年9月份日照时数210.9 h(济南)、238.3 h(德州)高于泰安、临沂9月份日照时数196.5和177.8 h。黄色素含量的高低总体和降雨量负相关,9月份谷子灌浆中后期较多的降雨对于黄色素在种子中的积累是不利的。温度、光照、降水等气象因子量的变化,光、热、水资源的时空分布对谷子生长产生重要影响。因此,试点之间气象因素的差异是黄色素含量差异的重要原因。
有研究[22]表明,适时收获有利于获得较高的黄色素含量,谷子过度成熟黄色素含量会急剧下降。本研究中,临沂、泰安试点播种到收获的天数超过100 d,临沂试点2017年达到105 d,济南、德州试点播种到收获都少于100 d,临沂、泰安试点谷子成熟后收获延迟可能也是这两个试点黄色素含量低的原因之一。因此,谷子成熟后及时收获对于获得较高的小米黄色素含量也是必要的,避免因为收获延迟影响小米的品质。不同地点的土壤质地、肥力水平可能对黄色素含量也有一定的影响,尚需要进一步研究。
4 结论
品种基因型是决定小米黄色素含量的最重要因素。不同年份、地点、品种及各因素互作对小米黄色素含量有较大的影响,谷子生育期平均气温、降雨量、光照时数等天气因素的变化及时空分布的差异是小米黄色素含量变化的重要原因;生育期平均气温较高,灌浆中后期光照充足有利于小米黄色素的积累。
[1] YANG X Y, WAN Z W, PERRY L, LU H Y, WANG Q, ZHAO C H, LI J, XIE F, YU J C, CUI T X, WANG T, LI M Q, GE Q S. Early millet use in northern China., 2012, 109(10): 3726-3730.
[2] BETTINGER R L, BARTON L, MORGAN C. The origins of food production in north China: A different kind of agricultural revolution.2010, 19(1): 9-21.
[3] LU H Y, ZHANG J P, LIU K B, WU N Q, LI Y M, ZHOU K S, YE M L, ZHANG T Y, ZHANG, H J, YANG X L, SHEN L C, XU D K, LI Q. Earliest domestication of common millet () in East Asia extended to 10,000 years ago., 2009, 106(18): 7367-7372.
[4] 王海棠, 尹卫平, 阳勇, 陈海涛. 小米黄色素的初步研究——化学成分及应用研究. 中国粮油学报, 2004, 19(3): 26-30.
WANG H T, YIN W P, YANG Y, CHEN H T. Primary study on millet yellow pigment-study on chemical constituents and application of millet yellow pigment., 2004, 19(3): 26-30. (in Chinese)
[5] SHEN R, YANG S P, ZHAO G H,SHEN Q, DIAO X M. Identification of carotenoids in foxtail millet () and the effects of cooking methods on carotenoid content.,2015, 61: 86-93.
[6] ZHANG L Z, LIU R H. Phenolic and carotenoid profiles and antiproliferative activity of foxtail millet., 2015, 174: 495-501.
[7] ROBEERTS R L, GREEN J, LEWIS B. Lutein and zeaxanthin in eye and skin health., 2009, 27: 195-201.
[8] 杨延兵, 管延安, 秦岭, 石慧, 王海莲, 张华文. 不同地区谷子小米黄色素含量与外观品质研究. 中国粮油学报, 2012, 27(1): 14-19.
YANG Y B, GUAN Y A, QIN L, SHI H, WANG H L, ZHANG H W. The studies on yellow pigment content and appearance quality of millet from different regions., 2012, 27(1): 14-19. (in Chinese)
[9] RAVEL C, DARDEVET M, LEENHADT F, BORDES J, JOSEPH J L, PERRETANT M R, EXBRAYAT F, PONCET C, BALFOURIER F, CHANLIAUD E, CHARMET G. Improving the yellow pigment content of bread wheat flour by selecting the three homoeologous copies of., 2013, 31: 87-99.
[10] WURTZWL E T, CUTTRISS A, VALLABHANENI R. Maize provitamin a carotenoids, current resources, and future metabolic engineering challenges., 2012, 3: 29.
[11] Fu Z, YAN J, ZHENG Y, WARBURTON M L, CROUCH J H, LI J S. Nucleotide diversity and molecular evolution of thegene incompared to some other grass species., 2010, 120(4): 709-720.
[12] KUHNEN S, LEMOS P M, CAMPESTRINI L H, OGLIARI J B, DIAS P F, MARASCHIN M. Carotenoid and anthocyanin contents of grains of Brazilian maize landraces., 2011, 91: 1548-1553.
[13] KEAN E G, BORDENAVE N, EJETA G, HAMAKER B R, FERRUZZI M G. Carotenoid bioaccessibility from whole grain and decorticated yellow endosperm sorghum porridge., 2011, 54: 450-459.
[14] Moros E E, Darnoko D, Cheryan M, Perkins E G, Jerrell J. Analysis of xanthophylls in corn by HPLC., 2002, 50(21): 5787-5790.
[15] DIGESÙ A M, PLATANI C, CATTIVELLI L, MANGINI G, BLANCO A. Genetic variability in yellow pigment components in cultivated and wild tetraploid wheats., 2009, 50: 210-218.
[16] HARJES C E, ROCHEFORD T R, BAI L, BRUTNELL T P, KANDIANIS C B, SOWINSKI S G, STAPLETON A E, VALLABHANENI R, WILLIAMS M, WURTZEL E T, YAN J, BUCKLER E S. Natural genetic variation in lycopene epsilon cyclase tapped for maize biofortification., 2008, 319: 330-333.
[17] YAN J, KANDIANIS C B, HARJES C E, BAI L, KIM E H, YANG X, SKINNER D J, FU Z, MITCHELL S, LI Q, FERNANDEZ M G, ZAHARIEVA M, BABU R, FU Y, PALACIOS N, LI J, DELLAPENNA D, BRUTNELL T, BUCKLER E S, WARBURTON M L, ROCHEFORD T. Rare genetic variation atcrtRB1 increases beta-carotene in maize grain., 2010, 42(4): 322-327.
[18] 张耀元, 路阳, 张彬, 禾璐, 刘龙龙, 王兴春, 李红英, 韩渊怀. 谷子类胡萝卜素生物合成途径SiLCYB基因与米色形成的关系, 分子植物育种, 2016, 14(6): 1341-1351.
ZHANG Y Y, LU Y, ZHANG BIN, HE L, LIU L L, WANG X C, LI H Y, HAN Y H. The relationship between the generelated to carotenoid synthesis and the colored formation of foxtail millet., 2016, 14(6): 1341-1351. (in Chinese)
[19] YANG Y B, JIA G Q, DENG L G, QIN L, CHEN E Y, CONG X J, ZOU R F; WANG H L; ZHANG H W, LIU B, GUAN Y A, DIAO X M. YIN Y P. Genetic variation of yellow pigment and its components in foxtail millet ((L.) P. Beauv.) from different eco-regions in China., 2017, 16: 2459-2469.
[20] LIU M X, ZHANG Z W, REN G X, ZHANG Q, WANG Y Y, PING L U. Evaluation of selenium and carotenoid concentrations of 200 foxtail millet accessions from China and their correlations with agronomic performance., 2016, 15(7: 1449-1457.
[21] 董倩楠, 刘艳芳, 程丽萍, 郭二虎, 陕永杰, 张美萍. 不同施肥条件对谷子β-胡萝卜素和叶黄素含量的影响, 核农学报, 2018, 32(5): 1003-1008.
DONG Q N, LIU Y F, CHENG L P, GUO E H, SHAN Y J, ZHANG M P. Effect of different fertilization conditions on the content ofβ-carotene and lutein in foxtail millet., 2018, 32(5): 1003-1008. (in Chinese)
[22] YANO A, TAKAKUSAGI M, OIKAWA K, NAKAJO S, SUGAWARA T. Xanthophyll levels in foxtail millet grains according to variety and harvesting time., 2016, 20: 136-143.
[23] Ning N, Yuan X Y, Dong S Q, Wen Y Y, Gao Z P, Guo M J, Guo P Y. Increasing selenium and yellow pigment concentrations in foxtail millet (l.) Grain with foliar application of selenite., 2016, 170(1): 245-252.
[24] 杨延兵, 张涵, 王润丰, 邓立刚, 秦岭, 陈二影, 管延安. 谷子籽粒小米黄色素含量的测定. 中国粮油学报, 2019, 34(3): 121-125.
YANG Y B, ZHANG H, WANG R F, DENG L G, QIN L, CHEN E Y, GUAN Y A. Determination of yellow pigment content in foxtail millet., 2019, 34(3): 121-125. (in Chinese)
[25] BENNETZEN J L, SCHMUTZ J, WANG H, PERCIFIELD R, HAWKINS J, PONTAROLI A C, ESTEP M, FENG L, VAUGHN J N, GRIMWOOD J, JENKINS J, BARRY K, LINDQUIST E, HELLSTEN U, DESHPANDE S, WANG X, WU X, MITROS T, TRIPLETT J, YANG X, YE C Y, MAURO-HERRERA M, WANG L, LI P, SHARMA M, SHARMA R, RONALD P C, PANAUD O, KELLOGG E A, BRUTNELL T P, DOUST A N, TUSKAN G A, ROKHSAR D, DEVOS K M. Reference genome sequence of the model plant., 2012, 30: 555-561.
[26] ZHANG G Y, LIU X, QUAN Z W, CHENG S F, XU X, PAN S K, XIE M, ZENG P, YUE Z, WANG W L, TAO Y, BIAN C, HAN C L, XIA Q J, PENG X H, CAO R, YANG X H, ZHAN D L, HU J C, ZHANG Y X, LI H N, LI H, LI N, WANG J Y, WANG C C, WANG R Y, GUO T, CAI Y J, LIU C Z, XIANG H T, SHI Q X, HUANG P, CHEN Q C, LI Y R, WANG J, ZHAO Z H. Genome sequence of foxtail millet () provides insights into grass evolution and biofuel potential., 2012, 30: 549-554.
[27] JIA G Q, HUANG X H, ZHI H, ZHAO Y, ZHAO Q, LI W J, CHAI Y, YANG L F, LIU K Y, LU H Y, ZHU C R, LU Y Q, ZHOU C C, FAN D L, WENG Q J, GUO Y L, HUANG T, ZHANG L, LU T T, FENG Q, HAO H F, LIU H K, LU P, ZHANG N, LI Y H, GUO E H, WANG S J, WANG S Y, LIU J R, ZHANG W F, CHEN G Q, ZHANG B J, LI W, WANG Y F, LI H Q, ZHAO B H, LI J Y, DIAO X M, HAN B. A haplotype map of genomic variations and genome-wide association studies of agronomic traits in foxtail millet ()., 2013, 45: 957-961.
[28] 杨延兵. 谷子黄色素及主要组分全基因组关联分析[D]. 泰安: 山东农业大学, 2017.
YANG Y B. Genome-wide association studies of yellow pigment and its components in foxtail millet ((L.) P. Beauv.)[D]. Taian: Shandong Agricultural University, 2017. (in Chinese)
[29] NDOLO V U, BETA T. Distribution of carotenoids in endosperm, germ, and aleurone fractions of cereal grain kernels., 2013, 139: 663-671.
[30] HOWITT C A, POGSON B J. Carotenoid accumulation and function in seeds and non-green tissues., 2006, 29(3): 435-445.
Yellow pigment contents in foxtail millet () under Diverse ecological conditions
YANG YanBing1, CHEN ErYing1, WANG RunFeng1, QIN Ling1, YIN XiuBo2, ZHANG HuiDi1, Li FeiFei1, Guan YanAn1
(1Institute of Crop, Shandong Academy of Agricultural Sciences/Shandong Engineering Laboratory for Featured Crop, Jinan 250100;2Shandong General Station of Agricultural Technology Extension, Jinan 250100)
【】Grains of(L.) are known as millet after dehulling. Yellow pigment content in grains is one of the crucial factors affecting the value of commodity as well as nutritional quality of foxtail millet. Investigating variations of yellow pigment content across ecological conditions would contribute to breeding and utilization of new foxtail millet varieties featured by superior qualities.【】 Eight newly released superior and distinctive foxtail millet cultivars from the summer- sowing region in North China were planted at five locations in two consecutive years (2016 and 2017). Yellow pigment contents of the foxtail millet cultivars were determined for each location. Analysis of Variance (ANOVA) was adopted to determine the significant levels of differences for yellow pigment contents of foxtail millet cultivars under ecological conditions. Correlation analysis was performed on between yellow pigment content, and air temperature, rainfall and sunshine duration.【】Growing years, cultivar genotypes, growing locations × growing years, cultivar genotypes × growing years, and growing years × cultivar genotypes × growing years showed highly significant effects on yellow pigment content (<0.01). Growing locations and growing locations×cultivar genotypes showed significant influences on yellow pigment content (<0.05). Cultivar genotypes, growing locations, and their interactions had high contribution rate (57.12%, 27.57% and 6.12%, respectively) to the variation of yellow pigment content. Generally, yellow pigment contents of cultivars planted in 2017 were higher than those in 2016. In terms of growing locations, yellow pigment contents of cultivars planted at Jinan, Dezhou and Jining were significantly higher than those at Taian and Linyi both 2016 and 2017 (<0.05). Yellow pigment content of the eight cultivars was averaged at 23.42 mg·kg-1and ranged from 18.56 mg·kg-1to 26.14 mg·kg-1. Among the cultivars, Zhonggu 2 had the highest yellow pigment content, while Jilügu 1 had the lowest. Moreover, there were no significant differences between Zhonggu 2, Jinuogu 2, Jigu 21, and Jigu19. However, yellow pigment contents of the above mentioned cultivars were higher than those of Yugu 18, Jigu 22, Jigu 20, and Jilügu 1. Furthermore, yellow pigment content was positively correlated with average temperature at seedling stage in June (=0.908,<0.01), average temperature at the middle and late grain filling stage (=0.798,<0.01), average temperature during growth period (=0.808,<0.01), sunshine hour in middle and later growth stages in September (=0.771,<0.01), and total sunshine hour of during growth period (= 0.769,<0.01).【】Cultivar genotypes, growing years, locations and their interactions significantly affected yellow pigment content in foxtail millet. Among these factors, cultivar genotype played a key role. Moreover, variations and spatial-temporal distribution differences in meteorological factors such as temperature, rainfall and sunshine hour in different years and locations were important for yellow pigment accumulation. The higher average temperature and more sunshine hours in the middle and late stages of grain filling were beneficial to the accumulation of yellow pigment. Cultivar genotypes are the most significant element influencing yellow pigment contents.
foxtail millet; yellow pigment content; ecological conditions; air temperature; rainfall; sunshine hour
10.3864/j.issn.0578-1752.2019.18.015

2019-05-28;
2019-06-27
山东省现代农业产业技术体系创新团队专项资金(SDAIT-15-03)、山东种业集团股份有限公司创新项目(ZYCX2016016)、山东省农业科学院创新工程(CXGC2018002)、现代农业产业技术体系专项(CARS-06)
杨延兵,E-mail:ybyang_666@163.com。
管延安,E-mail:yguan65@163.com
(责任编辑 李莉)
