水稻矮缩病毒对黑尾叶蝉生长发育和繁殖的影响
2019-10-10王前进叶恭银
王前进,方 琦 ,叶恭银
(1.洛阳师范学院生命科学学院,河南 洛阳 471000;2.浙江大学昆虫科学研究所/水稻生物学国家重点实验室/农业农村部作物病虫分子生物学重点实验室,浙江 杭州 310058)
【研究意义】水稻矮缩病毒(Rice dwarf virus,RDV)是水稻普通矮缩病(rice dwarf disease)的病原,属于呼肠孤病毒科(Reoviridae)植物呼肠孤病毒属(Phytoreovirus)[1]。该病害在水稻整个生长发育期内均可发生,严重时可造成水稻大面积减产,甚至绝收[2-3]。黑尾叶蝉Nephotettix cincticeps(Uhler)是水稻上重要的害虫,在我国大部分稻区都有为害,不仅可以直接刺吸水稻汁液造成危害,还可作为媒介昆虫以持久性方式传播水稻普通矮缩病、黄矮病和黄萎病等多种病毒病,间接地给水稻产业造成了更为严重的损失[4-6]。媒介昆虫是病毒病传播的重要途径,病毒病的传播依赖于特定的媒介昆虫,虫传病毒病害的防控不同于真菌与细菌等病害的防治,既要控制病毒本身,又要通过控制介体昆虫来控制病毒病害的蔓延与流行[7]。长期进化过程中,病毒-昆虫-寄主植物三者间形成了复杂而特异的互作关系,这种协同进化关系不仅是生态学、进化生物学等理论学科的重要研究内容,同时也是病害防控措施制定的理论基础[8]。因此,探明水稻矮缩病毒对黑尾叶蝉的生长发育和种群增长情况等的影响,对于害虫种群治理和水稻病毒病可持续控制具有重要意义。
【前人研究进展】近年来,以病毒-介体昆虫-寄主植物作为研究对象,从生态学水平、分子水平等解析三者复杂的互作关系取得了诸多重要进展[7]。病毒对介体昆虫的影响分为直接影响和间接影响两个方面,直接影响是病毒在媒介昆虫体内滞留对昆虫生物学特性的影响[9-10];间接影响是病毒侵染寄主植物后会引起植物的光合作用、次生代谢、营养成分、信号途径等发生变化,继而影响介体昆虫的生物学特性,最终会影响介体对于病毒的传播[8]。这种由寄主植物介导的三者互作关系是决定昆虫种群增长和田间病害流行的重要因素[11]。水稻矮缩病毒对其重要传播介体黑尾叶蝉的直接和间接影响报道较少。有学者发现相对于不携带水稻矮缩病毒的叶蝉,携带水稻矮缩病毒的黑尾叶蝉不育率增高,生殖力下降,推断水稻矮缩病毒对黑尾叶蝉的直接影响是不利的[12-13]。但黑尾叶蝉取食感染水稻矮缩病毒的稻株后,产卵量、寿命、存活率、显著高于对照[14],种群增长更快[15],推断水稻矮缩病毒对黑尾叶蝉的间接影响是有利的。但不同水稻品种间影响结果是否一致有待进一步验证。【本研究切入点】以籼稻TN1品种为试验材料,通过室内观测黑尾叶蝉取食健康与感病水稻后的生长发育、种群增长情况,并组建实验种群生命表,分析比较不同处理对黑尾叶蝉生长发育、种群增长等方面的影响。【拟解决的关键问题】明确水稻矮缩病毒对黑尾叶蝉生长发育及繁殖等生物学特征的影响,为分析病毒与介体昆虫“双重”为害提供佐证。
1 材料与方法
1.1 试验材料
健康昆虫种群:无毒黑尾叶蝉第一代于2015年8月采自浙江大学紫金港校区西区试验农场(杭州),在室内盆栽TN1水稻上饲养3代以上供试验。
带毒昆虫种群:取若干无毒低龄黑尾叶蝉若虫在感病TN1水稻上饲毒2 d后转移至健康TN1水稻上度过约1周的传毒潜伏期。随后,将叶蝉单头分别接到装有TN1幼苗(10 d)的大试管内(长25 cm,直径3 cm)让其取食传毒。2 d后将水稻苗取出并换入新的健康水稻苗,换出的幼苗移栽到人工气候室内培养。约15 d后观察水稻若有明显病症[16]并经过RT-PCR检测确认感染水稻矮缩病毒[12],对应的传毒叶蝉即为带毒叶蝉,收集并用感病的TN1水稻饲养,建立带毒种群3代以上供试验。
健康水稻:籼稻TN1,在人工气候箱内浸种培养10 d后移栽到人工气候室内,移栽后30 d的稻苗作为健康处理供试验。
感病水稻:方法与建立带毒种群相似,从带毒叶蝉种群中吸取4~5龄若虫接入装有TN1幼苗(10 d)的大试管中让其取食传毒,2 d后将水稻苗取出移栽到人工气候室内培养,30 d后选取症状明显的感病水稻供试验。
供试水稻均以木村B营养液[17]进行培养,控温控光控湿智能人工气候室培养条件为:温度27(±1)℃、光周期(明∶暗)14 h∶10 h、光强度3 500~4 000 Lx、相对湿度75(±5)%。
1.2 黑尾叶蝉生长发育参数和生命表的构建
将供试水稻移入装有木村B营养液[17]的自制养虫装置内(图1),接入无毒的初孵若虫,每装置1头。试验设置健康与感病水稻2个处理,每处理60个重复。逐日记录若虫的存活和蜕皮情况,至若虫羽化为止。待若虫羽化后称量成虫鲜重并记录。
收集健康与感病水稻上的黑尾叶蝉成虫,将其按雌雄比1∶1进行配对,置于养虫装置内相应处理的稻苗上。逐日更换新鲜稻苗,更换下的水稻苗在解剖镜下统计卵粒数,同时记录叶蝉存活情况,若雄虫死亡,则补充新的雄虫,直到雌虫死亡。
生命表参数根据以下公式计算:种群净增殖率R0=∑lxmx,世代平均历期T=∑lxmx/R0,内禀增长率rm=(lnR0)/T,周限增长率λ=erm,种群加倍时间td=ln2/rm。其中,x为发育时间,lx为年龄特征存活率,表示任一个体在x期间得以存活的概率;mx为年龄特征繁殖力,表示在x期间平均每头雌虫日产卵量。

图1 黑尾叶蝉的饲养装置及感病、健康水稻叶片Fig.1 Experimental device for Nephotettix Cincticeps,RDV-infected rice leaf and healthy rice leaf
1.3 黑尾叶蝉种群增长情况
将羽化24 h内的健康黑尾叶蝉成虫经饥饿处理6 h后,接入养虫笼内(50 cm×50 cm×50 cm)。每个养虫笼接入一对雌雄虫。接虫后,每月定时调查成虫数量。每周定期往养虫笼内添加新鲜水稻,待衰老水稻自然干枯后取出。试验设置健康与感病水稻2个处理,每个处理3次有效重复(雌虫至少存活20 d的处理作为有效重复,若雄虫死亡,则补充新的雄虫)。
1.4 数据分析
黑尾叶蝉在健康与感病2个处理水稻上的若虫期存活率采用SPSS(version 16.0)中的Kaplan-Meier法进行分析[18],若虫发育历期、寿命、鲜重、产卵量及种群数量等采用t测验进行分析,生命表参数采用Maia等[19]编写的SAS程序进行显著性分析。
2 结果与分析
2.1 水稻矮缩病毒对黑尾叶蝉生长繁殖的影响
黑尾叶蝉取食健康水稻时,13~31 d若虫全部孵化为成虫,期间若虫期存活率为70.00%。而黑尾叶蝉取食感病水稻时,若虫期存活率在28 d全部孵化为成虫时降到66.67%。两者进行数据分析后得出P= 0.843,未达显著水平(图2)。

图2 水稻矮缩病毒对黑尾叶蝉若虫期存活率的影响Fig.2 Effect of RDV on nymph survival rate of Nephotettix Cincticeps
黑尾叶蝉取食感病水稻时雄性、雌性若虫历期分别为21.29 、22.57 d,与取食健康水稻相比均显著缩短(雄性P= 0.034;雌性P= 0.001)。取食感病水稻发育而成的叶蝉雄性、雌性成虫鲜重均略有增加,但未达显著差异水平(雄性P= 0.588;雌性P= 0.907)。取食感病与健康水稻的雄性、雌性叶蝉成虫寿命则表现出较大差异:健康水稻上雄性叶蝉寿命为28.70 d,感病水稻上的雄性叶蝉寿命显著缩短为14.42 d(P<0.001);健康水稻上雌性叶蝉寿命为24.68 d,感病水稻上的雌性叶蝉寿命略有延长,但未达显著差异水平(28.48 d,P= 0.183);黑尾叶蝉取食感病水稻后产卵量增加为205.62粒,相比健康水稻上略有增加,但未达显著差异水平(P=0.251,表1)。

表1 水稻矮缩病毒对黑尾叶蝉生物学参数的影响Table 1 Effects of RDV on biological parameters of Nephotettix Cincticeps
2.2 水稻矮缩病毒对黑尾叶蝉实验种群生命表参数的间接影响
黑尾叶蝉取食感病水稻时,成虫的日产卵量很快达到高峰(35 d时达14.05粒/雌),以后逐渐下降;而取食健康水稻时,成虫的日产卵量是逐渐达到产卵最高峰的,在36 d时达到最大值11.19粒/雌。黑尾叶蝉取食感病与健康水稻时的年龄特征存活率(lx)具有相似的趋势(图3)。
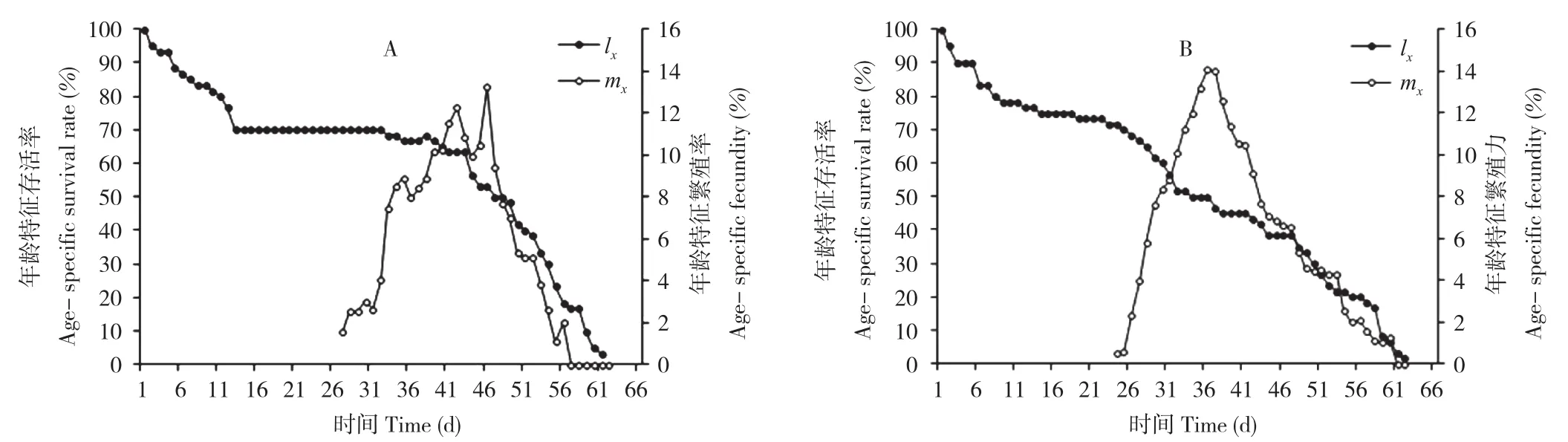
图3 水稻矮缩病毒对黑尾叶蝉年龄特征存活率(lx)和年龄特征繁殖力(mx)的影响Fig.3 Effects of RDV on age-specific survival rate (lx) and age-specific fecundity (mx) of Nephotettix cincticeps
根据黑尾叶蝉在健康和感病2个处理水稻上取食时各发育阶段的存活率和成虫的繁殖力,组建实验种群生命表,并计算出黑尾叶蝉的生命表参数(表2)。结果表明黑尾叶蝉取食感病水稻时净生殖率为138.29,显著高于取食健康水稻时的97.21;而2个处理水稻上内禀增长率、周限增长率、平均世代历期、种群加倍时间等4个生命表群参数间无显著差异。

表2 水稻矮缩病毒对黑尾叶蝉实验种群生命表参数的影响Table 2 Effects of RDV on life table parameters of tested population of Nephotettix Cincticeps
2.3 水稻矮缩病毒对黑尾叶蝉种群增长的影响
试验结果(表3)表明,随着调查时间的延长,2个处理水稻上成虫数量逐渐增大,前期增速较快,后增速逐渐放缓。接虫后3、4、5、6个月感病水稻上总的成虫数量分别为213.33、358.67、448.03、525.63头,健康水稻上相应的数据为136.33、235.71、292.34、351.67头,同一调查时间2个处理之间均达到显著差异水平(P= 0.032;P= 0.040;P= 0.032;P= 0.043)。说明黑尾叶蝉取食水稻矮缩病毒侵染的水稻后有利于种群的增长。
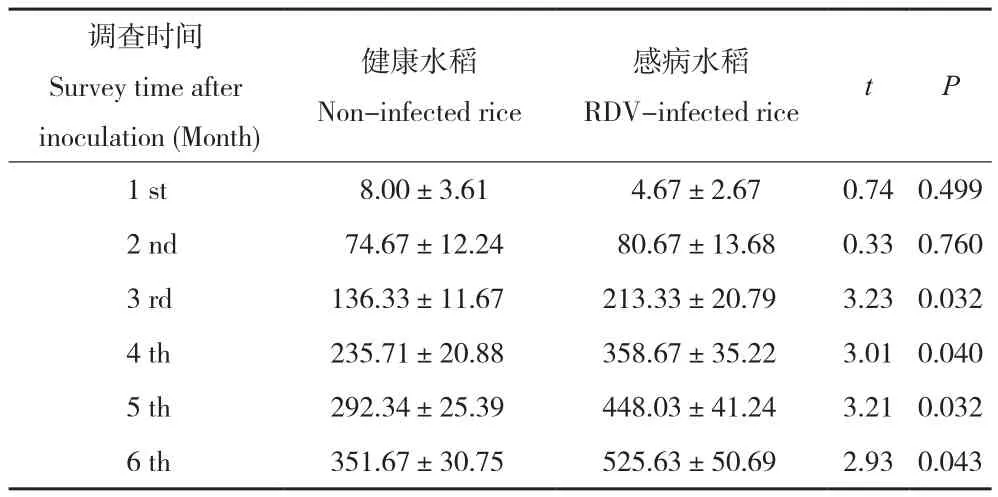
表3 水稻矮缩病毒对黑尾叶蝉种群增长情况的影响Table 3 Effects of RDV on population growth of Nephotettix Cincticeps
3 讨论
Wang等研究发现黑尾叶蝉取食感病水稻后其产卵量显著提高[14],而本试验中发现产卵量略有增加,但未达显著差异水平。以粳稻品种秀水11为供试水稻,研究发现叶蝉取食感病水稻时雌若虫发育历期显著缩短,雄成虫寿命显著延长[15],而本研究中发现雌雄若虫发育历期和雄性叶蝉寿命均显著缩短。这些结果的差异推测是水稻品种不同所致,不同水稻品种对水稻矮缩病毒的抗性差异较大[20],从而造成叶蝉的取食或营养利用有所不同。本研究发现黑尾叶蝉取食感病水稻时死亡率较低、产卵量较大,内禀增长力、净增殖率和周限增长率均有所提高,种群加倍时间缩短,且饲喂感病水稻时的叶蝉种群增长更快,种群数量更大,推断水稻矮缩病毒对黑尾叶蝉产生有利的间接影响,感病水稻更适合黑尾叶蝉种群的生长发育及繁殖。
植物病毒病传播过程涉及到昆虫、病毒、寄主植物三者的互作。目前有关植物病毒对媒介昆虫的影响报道较多,其中一些还对机理进行了探索。如小西葫芦黄花叶病毒(Zucchini yellow mosaic virus)造成介体棉蚜(Aphis gossypii)的寿命延长,生殖力显著提高,推断与感病植株汁液中氨基酸含量提高及糖分含量降低有关[21]。感染非洲木薯花叶病毒病毒(African cassava mosaec virus)的植株上烟粉虱(Bemisia tabaci)种群增长更快,推断与感病植株中氨基酸含量增高有关[22]。Figbig 等发现麦长管蚜(Sitobion avenae)在感染大麦黄矮病毒(Barley yellow dwarf virus)的大麦上内禀增长率rm显著降低,测定发现感病植株汁液氨基酸的含量明显降低,且对蚜虫蜜露的分析结果表明,蚜虫在感病植株上取食时对氨基酸的利用率较低,吸收的氨基酸含量相对较少[23]。另外,也有研究表明昆虫在感病寄主植物上适合度的提高与植物防御途径的改变有关,双生病毒能抑制寄主植物茉莉酸途径的关键转录因子,从而调控萜烯类化合物,吸引烟粉虱(Bemisia tabaci)的取食,促进双生病毒与烟粉虱建立互惠关系[24-25]。植物病毒能抑制植物茉莉酸抗虫信号途径,可以大大提高介体昆虫的适应度,形成了病-虫的互惠共生关系[26]。黑尾叶蝉是刺吸式口器昆虫,依赖通过口针吸食寄主植物汁液存活,而水稻感染水稻矮缩病毒后导致汁液成分的改变可能对叶蝉的取食或营养利用造成影响,且病毒的入侵也有可能导致植物防御途径的变化,从而导致其在感病水稻上的寿命、存活等生物学特性的变化。但水稻感染水稻矮缩病毒后导致汁液成分如何改变,植物防御途径如何相关,这些改变又如何影响黑尾叶蝉生物学特性的变化有待进一步研究阐明。
4 结论
本研究结果表明,水稻矮缩病毒(Rice dwarf virus)会促进黑尾叶蝉若虫的生长发育、加快种群增长速度,表明感病水稻更适合黑尾叶蝉种群的生长发育及繁殖。综上所述,我们认为水稻矮缩病毒对黑尾叶蝉会产生有利的间接影响。这种有利的间接影响可能是由于水稻感染水稻矮缩病毒后导致汁液成分的改变,或植物防御途径的变化等,继而对叶蝉的取食或营养利用造成改变的结果。在农业生产上,进一步加强抗性品种的选育、加强虫情病情的监测对于水稻普通矮缩病的防治至关重要。
