柳杉全同胞家系生长和材性的遗传变异及 优良家系选择
2019-10-10莫家兴翁怀峰曹玉婷戴丽华黄信金欧阳磊
莫家兴,华 慧,翁怀峰,曹玉婷,戴丽华,黄信金,欧阳磊,徐 进
(1.南京林业大学 林学院,江苏 南京 210037;2.南京林业大学 南方现代林业协同创新中心,江苏 南京 210037;3.福建省霞浦国有林场,福建 霞浦 355100;4.福建省林业科学研究院,福建 福州 350012)
林木产量和质量是林木遗传改良重要研究内容,分析和掌握林木生长与材性性状变异对提高林地生产力和提升经济效益具有重要意义。木材利用效益受生长与材性多性状综合影响,生长性状决定了木材产量,而材性性状则决定了木材质量[1]。木材质量高低与木材的硬度、弹性、抗压缩及抗弯及纤维(管胞)的长度、宽度等都有着密切的关系,一般来说,遗传变异影响木材性状变异[2],木材密度影响木材强度[3],木材密度是反映木材力学性能的最佳预测指标[4,5],而纤维(管胞)长度是评价纸浆特性和造纸质量的重要指标之一。
林木生长与材性性状大多由多基因控制,但一些报道发现具有高遗传力的性状[6-9]受到相对较少基因控制[3],这些高遗传力性状对于优良家系选择具有十分重要的意义。有研究表明,林木的生长与材性性状之间没有相关性,表现为独立遗传,但也有少数研究认为林木生长与材性性状间存在较弱的正相关或负相关[10-12]。
柳杉属Cryptomeria共2 种:柳杉Cryptomeria fortunei和日本柳杉C.japonica。柳杉是我国南方主要的针叶用材树种,具有生长快、寿命长、用途广等特点,是我国长江流域和南方各省主要造林树种之一。其木质偏轻但强度大,具有易加工、吸附能力强、热传导率低、触感好等一系列特点,已被广泛用于板材和建筑生产中。对柳杉家系和无性系生长及材性曾有一些研究,但主要是从生长性状方面评价家系[2,13-14]。通过对30年生柳杉全同胞家系的杂交子代林的生长性状(树高(height,h)、 胸 径(diameter at breast height,dbh) 和 材 积(volume,v))和材性性状(比重(specific gravity,sg)、心材率(hardwood ratio,rh)、偏心率(eccentricity,et)、树皮率(bark ratio,rb)、管胞长度(tracheid length,lt)、管胞宽度(tracheid width,wt)和管胞长宽比(length-width ratio of tracheid,rt))进行测定和遗传变异分析,综合评价柳杉40 个家系,选育出生长快、木材材性好的优良家系,为柳杉生长及材性改良提供依据。
1 材料与方法
1.1 材料
1.1.1 试验点基本情况
试验林位于福建省霞浦国有林场杨梅岭工区,海拔157 m,立地等级Ⅲ类。属于亚热带季风湿润气候区,年平均气温16~19 ℃,春多雨水,夏多台风,冬暖夏凉,霜雪少见,季风特点明显。
试验林于1984年定植,1年生实生苗种植,单株小区,重复12 次,造林设计采用完全随机区组设计,造林密度2 940 株/hm2。
1.1.2 材料来源
试验林地调查及木芯采集于2015年1月进行。根据试验林地保留情况,选取了其中保留较好的40 个全同胞家系,每个家系分别选5 株平均木,共200 株,家系组合见表1。30#家系(霞浦7 号×霞浦7 号)为对照家系。
1.2 方法
1.2.1 生长性状测定
常规方法测量树高、胸径并按照以下公式计算材积:

1.2.2 材性性状测定
1)木芯取样 用林木生长锥(瑞典Haglof,φ5.15 mm)在树木上坡方位树高1.3 m 处采集通过树皮和髓心的一条完整无疵的木芯,置于长形塑料管中保存,进行编号和记录,将所采集样品带回实验室进行材性相关性状测定。
2)心材率、偏心率和树皮率计算
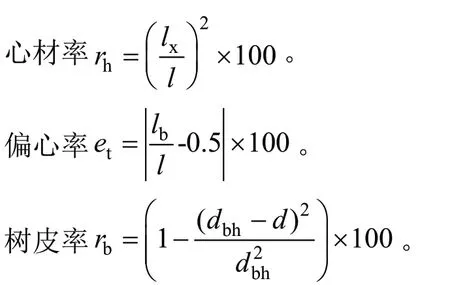
lx为木芯芯材部分长度(cm),l表示木芯长度(cm),lb代表半边木芯长度(cm),dbh表示胸径(cm),d表示树皮厚度(cm)。
3)木材比重、管胞长度、管胞宽度测定
采用Smith[15]方法测定木材比重。管胞形态性状测定采用过氧化氢—冰醋酸离析法。解剖针挑取部分管胞置于载玻片上,加数滴0.1%番红溶液后,加盖盖玻片后完成制片过程。将制片置于ZEISS Axioscope.A1 显微镜下观察,每个家系随机挑选30 个形态完整的管胞测量长度和宽度,并计算管胞长宽比(管胞长度/管胞宽度)。
4)其他相关参数计算
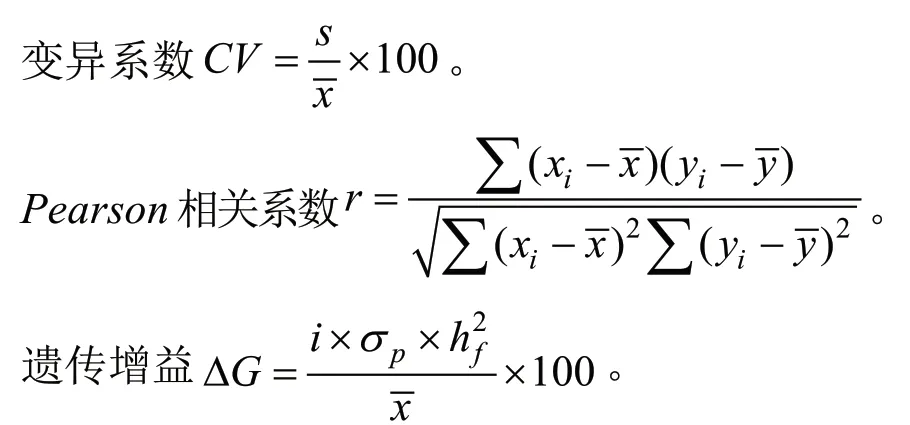
s表示性状标准差,CV表示性状变异系数,r表示相关系数,xi表示各家系性状平均值,为家系性状平均值,yi表示各家系另一性状平均值,表示家系另一性状平均值,σp代表亲本群体标准差,i代表选择强度,h2f为家系遗传力。
1.2.3 家系评价
以简单指数法进行家系选择,
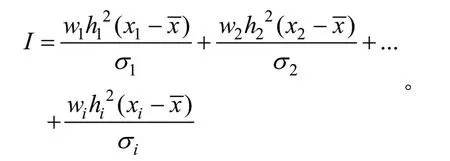
其中:I表示指数值,1、2…i表示参与选择的性状,w表示性状的加权系数,x表示各家系各性状的平均值,是总的性状平均值,h2是性状的遗传力,σ为性状的标准差。
不同入选率下的选择强度及遗传增益的估算参照王明庥[16]的方法。
2 结果与分析
2.1 柳杉不同家系间生长及材性差异分析
对柳杉杂交子代林林分进行调查,发现不同杂交组合的子代在生长量上和外形表现上均有较大差异,对40 个家系生长量进行统计可知,40个家系树高总平均值为12.2 m,其中15#家系最高,达13.6 m,而8#家系最小,仅有10.6 m;胸径平均为16.7 cm,其中10#和34#家系最大,为19.8 cm,22#家系最小,为13.3 cm;材积方面,平均值为单株0.139 4 m3,最大的为15#家系,单株达0.213 6 m3,最小的22#家系为0.083 0 m3。另外树高、胸径和材积变异系数均值分别为9.9%、13.8%和33.052 0%,材积的变异系数最大,其中2#家系变异系数最大,为57.611 6%,说明不同柳杉家系的生长量上差异性较大。
木材比重平均值为0.379 g/cm3,最大的34#家系木材比重达到0.437 g/cm3,最小的为28#家系比重为0.349 g/cm3;心材率平均值为40.0%,最大的49.2%为19#家系,最小的32.28%为14#家系。偏心率平均值为3.5%,最大的12.20%为13#家系,最小的0.56%为20#家系;另外树皮率平均值为3.3%,最厚的5.0%为25#家系,最薄的2.5%为3#家系和7#家系;木材比重、心材率、偏心率和树皮率的变异系数分别为7.73%,18.9%,82.4%和25.1%。
40 个家系的管胞长度平均值为2 556.21 μm,2#家系管胞长度最大,达到2 782.21 μm,32#家系管胞长度最小,仅有2 168.22 μm;管胞宽度平均值为33.74 μm,5#家系管胞宽度最大,达到37.99 μm,12# 家系管胞宽度最小,仅有31.33 μm;管胞长宽比平均值为78.22,其中2#家系管胞长宽比最大,达到88.69,32#家系管胞长宽比最小,仅有62.76。管胞长度、管胞宽度及管胞长宽比变异系数分别为6.49%、10.03%和11.42%,其中管胞长度变异系数最大的为5#家系,达17.43%;管胞宽度变异系数最大的为28#家系,达23.93%;管胞长宽比变异系数最大的为5#家系,达28.96%。
对柳杉不同家系生长和材性性状进行方差分析(表2),可知40 个家系间的树高、胸径、树皮率、管胞长度、管胞宽度和管胞长宽比均达到了差异极显著水平,材积和比重性状达到显著差异水平,而木材心材率和偏心率则差异不显著。

表2 家系间生长与材性性状方差分析及遗传力估算†Table2 Analysis of variance of growth and wood properties as well as calculation of heritability of families
2.2 柳杉不同家系生长及材性变异分析
优良家系选择因目的不同,所采取的选择方式也有不同,在单因素选择方面,家系各性状的遗传力、平均值和变异系数可作为其选择的重要依据。遗传力的大小反映特定性状遗传能力的大小;平均值大小决定了各家系总体在该性状数量上的高低,而变异系数则反映性状所具有的进一步遗传变异的潜力,同时也存在遗传选择的风险。
由表2可知,作为生长性状的树高和胸径具有中等偏上的单株遗传力水平(0.64 和0.66),同时具有中等水平的家系遗传力(0.49 和0.50),树皮率也有类似的情况(单株遗传力为0.59,家系遗传力为0.46)。比重具有中等偏上的单株遗传力(0.67)和中等家系遗传力(0.50);而管胞长度、管胞宽度和管胞长宽比这类材性性状则具有较低水平的单株遗传力(0.26、0.18 和0.24)和较高的家系遗传力(0.86、0.81 和0.85);而心材率和偏心率则具有较低水平遗传力(均小于0.2)。
以各家系的生长及材性性状的平均值与其变异系数绘制散点图(图1),评价各家系不同性状自身的优势及改良的潜力。图1中每个点代表一个家系,家系编号在散点旁标注,横坐标示各性状家系平均值,纵坐标示各性状变异系数值;坐标内十字线的纵线和横线分别表示家系的性状平均值和变异系数平均值。
在各家系树高均值和变异系数方面,由图1-A可知,均值最大的为15#家系,达13.6 m,其变异系数为6.6%,为一个变异幅度较低的情况。而均值较小的几个家系如3#、4#和9#,均值分别为10.8 m、10.9 m 和10.7 m,其变异系数却在一个相对比较高的水平,分别为12.9%、13.5%和13.5%。而变异系数最大的家系6#,达到了16.9%,其均值则在各家系中处于中间水平。
图1-B 中可知,在胸径方面,均值最大家系为10#和34#,均为19.8cm,而其变异系数分别为11.9%和14.4%,处于相对中间水平。均值最小的22#其变异系数也处于中间水平,为12.7%。均值和变异系数均较大的家系为2#,分别是19.6cm和26.9%。
比重方面,均值最大的家系是34#,达到0.437 g/cm3,远高于其他家系,其变异系数则处于平均水平,为8.08%。变异系数最高的家系28#,其均值最低,仅为0.349 g/cm3。而12#则具有均值和变异系数都较高的情况,分别是0.414 g/cm3和11.06%。
单株材积性状中均值最大者为15#,为 0.213 6 m3,其变异系数也达到了54.8%,属于一种高程度变异系数。其他家系中均值和变异系数都较高的有2#、12#和17#。
心材率方面,均值最大的19#,变异系数为14.1%,处于中下水平。而均值和变异系数均较高的家系则包括1#、4#和18#,其中以18#最突出。
偏心率与其他性状有所不同,其数值越小,则树木主干越通直,心材部分生长越均匀,对家系选择越为有利。由1-F 可知,在偏心率方面,均值最小的20#,偏心率为0.6%,其变异系数也最小,仅为29.9%。而25#、26#和28#则同时具有较小的均值和较大的变异系数。
树皮率也和偏心率相似,数值越小,树木主干可利用部分则越大,符合一般家系选择方向。由图1-G 可知,5#、20#和34#为具有较小的均值和较大的变异系数的家系。

图1 各家系各性状平均值与变异系数散点图(A~J)Fig.1 Scatter diagram of mean value and coefficient of variation of families (A-J)
由图1-H 可以看出,管胞长度方面,具有最大均值的是2#和4#,其值分别为2 782.21 μm 和2 763.53 μm,变异系数仅分别为1.29%和3.95%。家系27#和33#则有着较大的均值和变异系数。

续图1 各家系各性状平均值与变异系数散点图(A~J)Continuation of fig.1 Scatter diagram of mean value and coefficient of variation of families (A-J)
管胞宽度越小,则管胞长宽比越大,这对造纸质量有着重要影响。由图1-I 可以看出,家系1#、14#、16#和36#同时具有较小管胞宽度和较大变异系数。
管胞长宽比方面,家系2#有最大的均值,达到了88.69,其变异系数仅为5.95%。家系27#则同时有着较高的管胞长宽比和变异系数。
2.3 柳杉生长与材性性状相关分析
将柳杉有关生长及材性性状进行Pearson 相关分析,结果见表3。柳杉树高与胸径、材积存在极显著正相关;胸径与材积、管胞长度、管胞长宽比存在显著水平正相关,材积与管胞长度、管胞长宽比也存在相同情况;心材率与偏心率存在极显著正相关,而与树皮率则有极显著负相关;管胞长度和宽度管胞长宽比存在极显著正相关;其他性状间则为不显著的正相关或负相关关系。
2.4 柳杉不同家系评价
由于柳杉优良家系选择主要用于板材和纸浆生产,因此比重和管胞大小(管胞长度或长宽比)成为考量优良家系选择的关键。各性状相关分析(表3)表明管胞尺寸(管胞长度或长宽比)与胸径存在极显著正相关关系,而胸径的大小也对材积起了决定性的作用,故选择胸径和比重作为简单指数选择法的选择性状,其权重比例确定为 w胸径:w比重=0.5:0.5。各家系选择指数值I值计算结果见表4。按10%入选率,选出I最高的4 个家系,分别是34#(7-3)、3#(6-2)、2#(7-5)和20#(20-10),其I值分别为0.680 1、0.473 5、0.392 1 和0.361 1。这4 个家系的子代生长快且木材性质较好,可作为选育对象加以开发利用。
根据评定结果,以不同的入选率,对柳杉不同家系进行选择,选择后的生长及材性性状的遗传增益见表5。
当入选率为50%时,即40 个家系中将有20个被去除,所保留的20 个家系为表7中前20 名家系,它们的平均胸径、材积、比重和管胞长度分别提高6.5%,12.5%,3.5%和5.3%。当入选率为10%,即40 个家系中将有36 个被去除,所保留的4 个家系为表7中前4 名家系,即34#,3#,2#,20#,胸径、材积、比重和管胞长度遗传增益分别为14.3%,27.5%,7.8%和11.6%。随着入选率的降低,入选的家系减少,选择强度加大,获得的遗传增益也增大。

表3 生长与材性性状间相关性分析†Table3 Pearson correlation coefficient of traits among growth and wood properties

表4 40 个家系经简单指数选择法I 值排名†Table4 Rank of I value of simple index selection method of 40 families

表5 不同入选率下柳杉生长及材性性状的遗传增益Table5 The genetic gain of growth and wood properties under different selection rates
3 结论与讨论
对多个用材树种生长量变异进行研究,发现它们普遍存在树高、胸径和材积等生长性状显著变异。黄寿先等对杉木半同胞家系生长遗传变异研究表明树高、胸径和材积在家系间存在显著差异[1]。王润辉对17年生和19年生杉木无性系林分生长和材性性状变异进行研究,结果表明杉木无性系间树高、胸径、单株材积存在极显著的遗传差异[17]。本研究中柳杉全同胞子代各家系的树高、胸径和材积变异系数均值分别为9.9%、13.8%和33.05%,其中材积变异系数最大,且各家系间树高、胸径存在极显著差异,单株材积存在显著差异,这种差异程度有益于将来柳杉在生长性状方面选择与改良相关工作的开展。
木材品质是林木改良重要内容之一,多个用材树种在材性改良方面已经开展过不少研究。通过对杉木无性系测定林的生长和材性变异研究认为心材率和木材基本密度存在极显著差异[17]。尚秀华等对50 个赤桉家系研究发现树高、单株材积、木材基本密度、纤维长度、纤维宽度等在家系间差异均达到极显著水平[18]。施季森等较为系统地研究了自由授粉、测交及双列杂交交配系统中杉木的生长和材性的遗传变异系统,生长和材性性状均表现出极显著差异,其中木材比重在家系间存在极显著差异,受到中等强度的遗传控制[19]。本研究发现柳杉不同家系间树皮率、管胞长度、管胞宽度和管胞长宽比均存在极显著差异,比重存在显著差异,表明这些性状具有较大改良潜力。
有研究表明柳杉木材的强度,抵抗顺纹压缩及抗弯能力均低于杉木[20]。长纤维(管胞)是木材性质利用的另一个重要方面,纤维(管胞)长度是影响纸浆特性和造纸质量的重要因素。因此,对柳杉木材的强度、抗压抗弯能力及管胞长度的选择是改良的重要方向。本研究表明柳杉家系间管胞长度、管胞宽度和管胞长宽比存在极显著差异,管胞长度的单株遗传力较低(0.26),但家系遗传力较高(0.86),胸径也与管胞长度、管胞长宽比存在显著正相关。因此,通过加强林地管理改善树木生长环境,可以在一定程度上得到更多生产上尺寸理想的管胞,提高木材质量。
关联遗传是研究复杂树木性状的一种非常有效方法[21,22]。材性性状与许多其它性状相关,被认为是复杂的性状特征。例如,木材硬度受密度、纤维素结晶度、微纤丝角等方面影响。在多性状综合评价时,可以参考性状间相关关系进行评判。此外,主成分分析、聚类分析也可以应用于选择优良家系(无性系)[23,24]。
本研究中材积与树高、胸径、管胞长度和管胞长宽比存在极显著或显著的正相关关系,因此,在木材生产量上选择材积为主要性状,同时它也正向影响管胞长度,而木材比重是影响木材质量的主要性状,进行综合评价时考虑材积和木材比重,可以找出优良家系,培育出柳杉优质高产子代。纤维素、半纤维素、木质素等物质是木材主要组成部分,前二者对纸浆造纸产品质量有重要影响,木质素含量则决定了木材性质,在今后研究中可以将这些指标纳入柳杉优良家系选择考虑范围。
