地肤的开花动态及果实形态发育特性
2019-10-08巴特衣古丽·沃木尔扎克魏岩
巴特衣古丽·沃木尔扎克 魏岩


摘要:本试验以分布于准噶尔盆地南缘的地肤为材料,对其开花物候、花部综合特征及果实发育特性进行观察研究。结果表明:地肤于6月中下旬现蕾并开花,8月末至9月初结束开花,花期为60~70 d,为持续开花模式。地肤穗状花序由下部向顶部开放,花期持续50~60 d;地肤单花开放时间持续11~15 d,花期分为雌性亚期和雄性亚期,其中雌性亚期10~14 d,雄性亚期0.5 d。 地肤果实于 8月上旬开始膨大,9月后干、鲜重不断增加,长、宽度不断增大,10月中旬成熟。
关键词:地肤;开花物候;雌性先熟;果实发育
中图分类号:S567.210.1 文献标识号:A 文章编号:1001-4942(2019)08-0041-05
Abstract Kochia scoparia distributed in the southern margin of Junggar Basin was chosen as material to study its flowering phenology, flower characteristics and fruit development characteristics. The results showed that the flower bud of K. scoparia appeared and began to bloom at mid-to-late June, then ended flowering in late August to early September. The flowering period was 60~70 days, showing a continuous flowering pattern. The flowering order of spica was from lower part to top, and the corresponding flowering period lasted for 50~60 days. The blooming time of single flower was 11~15 days. Its flowering stage was divided into female subphase and male subphase, in which, the female subphase was 10~14 days, and male subphase was 0.5 days. The fruit of K. scoparia began to expand at the beginning of August, and its dry weight,fresh weight,length and width continuously increased after September, and it matured in mid-October.
Keywords Kochia scoparia; Flowering phenology; Protogyny; Fruit development
開花物候是植物生殖生态学研究的重要内容之一[1],也是植物生活史中的一个重要性状,通过传粉、种子扩散以及种子萌发和幼苗定居而影响植物个体的适合度[2,3],可在许多层面表现,如植物群落、科、属、种、个体、花序和单花,一定程度上反映了植物遗传和环境因素与开花模式之间的关系[4,5]。开花物候包括花部综合特征、花开放式样、花部各性器官的寿命[6]。对于开花物候的研究一般包括开花时间、开花峰值期和开花持续时间等[7]。作为植物重要的适合度因子,物种间开花物候的差异能够影响群落的结构、功能和多样性,并影响群落水平上的进化过程[8,9] 。
果实是被子植物特有的生殖器官,它的发育主要包括两个阶段,即果实膨大和果实成熟[10]。果实发育过程中,形状、大小、重量等均会发生一定变化。地肤(Kochia scoparia)是藜科(Chenopodiaceae)地肤属一年生草本植物,广泛分布于欧亚大陆的干旱和半干旱地区,是中亚干旱地区适口性好、营养价值高的牧草植物资源,适应性较广,并具有食用、药用、观赏等多种功能。新疆植物志[11]记载地肤属植物除木地肤(Kochia prostrata)是小半灌木外,其它均为一年生草本植物,如尖翅地肤(Kochia odontoptera)、伊朗地肤(Kochia iranica)、全翅地肤(Kochia krylovii)、毛花地肤(Kochia laniflora)、黑翅地肤(Kochia melanoptera)、地肤(K. scoparia)、碱地肤(Kochia scoparia var. sieversiana)。其中木地肤(K.prostrata)有2个变种,分别为灰毛木地肤(Kochia prostrate var. canescens) 和密毛木地肤(Kochia prostrata var. villosissima)。
前人关于藜科植物白滨藜(Atriplex cana)、驼绒藜(Ceratoides latens)[12]、小蓬(Nanophyton erinaceum)[13]、亚麻叶碱蓬(Suaeda linifolia)[14]、木地肤(K. prostrata)[15]等的开花物候和结实特性研究已见报道,但尚未在地肤上开展此类研究。本试验以分布于准噶尔盆地南缘的地肤为材料,对其开花物候、开花动态以及果实发育特征进行观察研究,旨在为生殖适应对策提供基础资料,同时为地肤的合理开发利用提供依据。
1 材料与方法
1.1 试验地概况及试验材料
试验于2018年5—11月在准噶尔盆地南缘进行。该区(43°80′99″ N,87°56′80″ E)春秋季湿润,夏季干旱,冬季长而寒冷,属于中温带荒漠地区,年降水量277 mm,年均温6.7℃,最高温度36.5℃,最低温度-9℃。
选取该区域自然生长的地肤健株为试材。
1.2 地肤开花物候观察
参照蔡丹红[12]、布海丽且姆·阿卜杜热合曼等[13]观察植物开花物候的方法进行地肤的野外观测,随机标记自然群体中生长良好和大小基本一致的植株,对个体、种群进行系统观测。蕾期到始花期开始每天观察1次。观察物候指标包括蕾期、始花期、开花高峰期、末花期、果实成熟过程。
个体水平上,始花期指第一朵花在个体上开放的时期;开花高峰期(盛花期):个体开花达到50%以上;终花期:无花开放。
种群水平上,25%的个体开花为始花期;50%的个体达到开花高峰期为种群开花高峰期;95%的植株开花结束为终花期[16]。
1.3 地肤花部综合特征及开花动态观察
在标记的地肤中随机选取10个穗状花序,每标记的花序中随机选2个花蕾,从蕾期到进入雌性期开始每半天观察1次,进入雄蕊期后每20 min观察1次,观察对象包括花各部分的形状、颜色和空间位置,花序开放顺序、持续时间,单花花朵开放过程中雄蕊和雄蕊的变化。
1.4 地肤果实发育阶段观察
随机标记20个生殖枝进行果实发育的观察,从幼果期(子房开始膨大)到果实成熟期每天观察1次。果实发育阶段,每隔2周随机采集20个果实(子房),用Nikon SMZ1000显微镜测量果实的长度和宽度,使用万分之一电子天平称量果实鲜重和干重。
1.5 数据处理
用Microsoft Excel 2010进行数据统计及制图,用SPSS 19.0统计软件进行数据方差及显著性分析。
2 结果与分析
2.1 地肤种群、个体的开花物候
经观察,地肤花期为6月下旬至8月下旬。6月中旬进入现蕾期(蕾期持续3~5 d),6月下旬进入始花期(始花期持续10~15 d ),7月中旬进入盛花期(约35 d),8月末至9月初花期结束,10月中下旬果实成熟。地肤种群花期为60~70 d;个体花期为50~60 d;单花水平上的花期达到11~15 d。地肤种群、个体的开花物候详见表1。
2.2 地肤花部综合特征及开花动态
经观察发现,地肤花序生于主枝和侧枝上部。主枝上开花顺序:下部→上部→顶部,同一生殖枝中下部的花序先开放,约3~7 d后上部的花序再开花。侧枝上的花序与主枝下部花序同步开花,顺序为基部→顶端。
地肤花属于典型的风媒花,花小、不鲜艳;花两性,常形成复穗状花序。1~3朵生于上部叶腋,花下有时有锈色长柔毛;花被近球形,淡绿色,无毛或先端稍有毛;雄蕊5枚,稀有4枚或6枚,花丝白色扁平,花药宽矩圆形、外伸,花药颜色有黄色、红色、红黄之间的过渡色;子房上位、卵形,花柱纤细,柱头2个、丝状。
地肤是自花授粉植物,花有雌雄异熟现象,雌性先熟,同一花朵的雌蕊在雄蕊散粉前10~12 d就已成熟,授粉时雌蕊大部分都枯萎。雌蕊根据柱头的形态变化分为4个阶段完成成熟,包括柱头露出、柱头展开、受粉、萎蔫,整个过程持续时间为10~14 d。柱头完成受粉后,子房迅速生长成饱满的球形,之后苞片裂开,原来卷曲的雄蕊从花被片伸出。此时剥掉花被片可以看到弯曲的花丝,花药紧紧排列在花被片下,开花过程进入雄性阶段。雄蕊一般在8∶ 00—11∶ 00露出,雄蕊露出约为30 min,花药被花丝推到柱头外面,花丝伸直托花药,花药开裂并散粉,散粉时间约为30 min,雄性期只持续0.5 d。由此可知,地肤开花分为雌性亚期和雄性亚期(图1)。
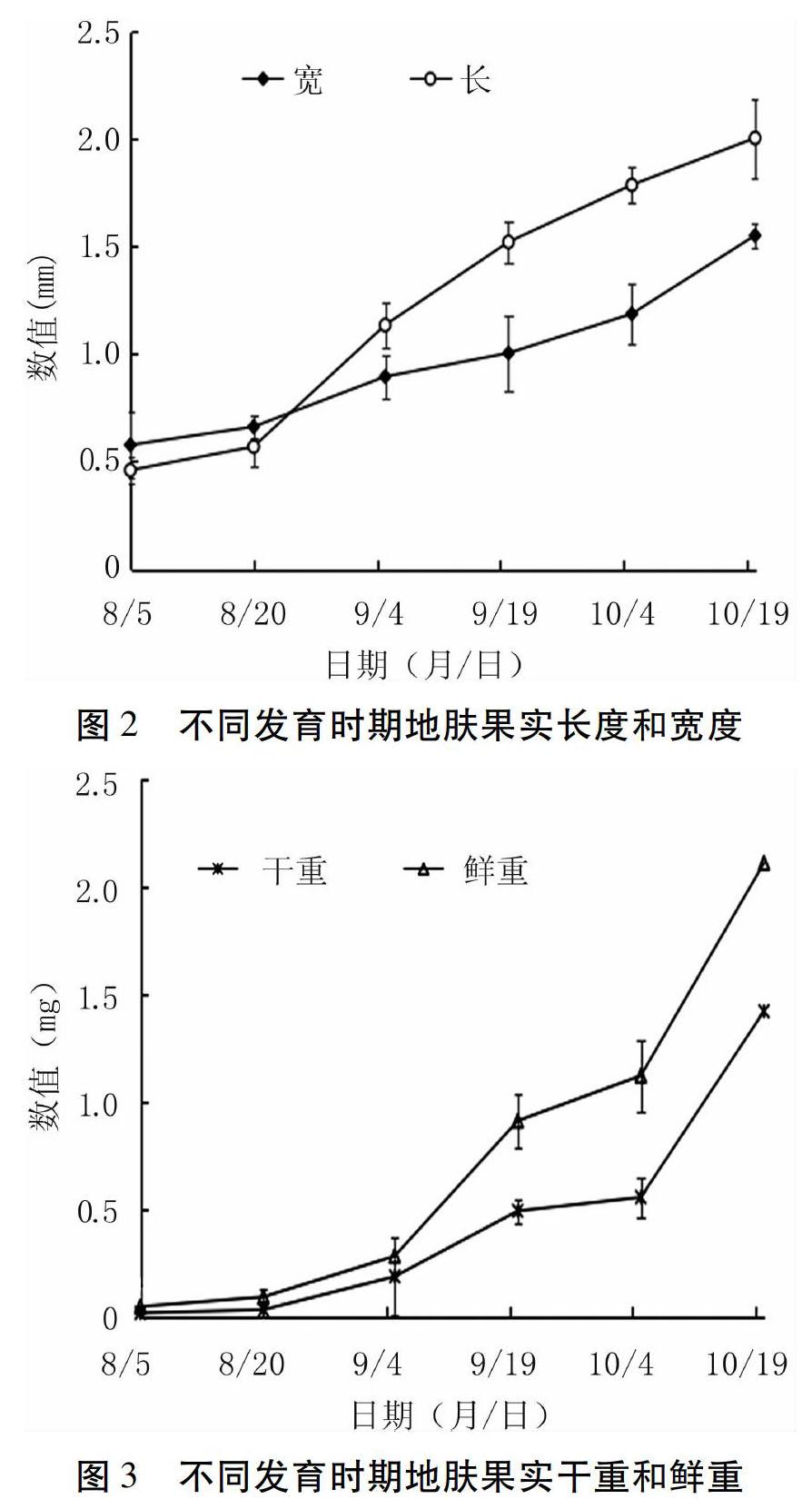
2.3 地肤果实发育
由图2、3可知,地肤果实在 8月上旬开始发育,此时干、鲜重和长、宽度变化不明显,9月4日时果实大小和重量开始发生显著变化,并随时间的延长,干、鲜重不断增加,长、宽度不断增长,至10月中下旬发育完成,达到成熟。而花被片在地膚开花后不脱落,并随着果实发育,背部向外突起形成翅状附属物。
3 讨论与结论
开花物候作为植物生殖物候的一个方面,涉及始花期、开花高峰期(盛花期)、开花持续时间等指标[17],其中始花期在植物功能属性研究中起着重要作用[18]。本试验在观察中发现,地肤于6月下旬进入始花期,7月中旬进入开花高峰期,该时期持续时间最长,约为35 d;地肤种群的花期达到60~70 d,个体的花期50~60 d,为持续开花模式。该结果与郭红超[15]关于木地肤开花进程的研究相似。
两性花中,为避免不同性器官功能相互干扰对花粉输入、输出产生影响,通常某一性器官先成熟行使其功能后,另一性器官再成熟,即雌雄异熟[19]。雌雄异熟被认为是植物用来减少自交和相互作用[20,21]的一种机制。本试验中,地肤单花花期为11~15 d,存在雌性先熟现象,雌性期较长,持续10~14 d,雄性期很短,持续0.5 d。雌性先熟在促进异型杂交方面更有效,因为雌性先熟植物的柱头在花药成熟前成熟,有利于优先接受异株植物花粉实现异型杂交[13]。
环境因子是制约荒漠植物生存的关键因素,水分和温度影响荒漠地区植物的开花物候。植物在长期的进化过程中表现出与环境相适应的特征,如藜科植物果实发育期较长,在湿度相对高的季节营养生长旺盛、储备养分,在夏季高温、湿度低、蒸发量大的时间段持续性开花,秋季温度降低时果实迅速发育成熟[22]。本试验中地肤夏季开花且花期较长,开花期间果实发育相对缓慢,9月上旬果实(种子)的长、宽度,干、鲜重明显增加,10月中下旬温度降低时种子快速发育成熟。地肤在秋季低温条件下果实迅速发育的机制与亚麻叶碱蓬等相似,主要是为了逃避荒漠地区夏季高温干旱的恶劣环境,也是在长期进化过程中适应荒漠气候、抵抗自然干扰的一种生殖保障。
参 考 文 献:
[1] Willson M F. Plant reproductive ecology[M].New York :John Wiley and Sons, 1983:80-85.
[2] Peuelas J, Filella I. Responses to a warming world[J]. Science, 2001, 294(5543):793-795.
[3] Wright S J, Calderon O. Phylogenetic patterns among tropical flowering phenologies[J]. Journal of Ecology, 1995, 83(6): 937-948.
[4] Davies T J, Wolkovich E M, Kraft N J B, et al. Phylogenetic conservatism in plant phenology[J].Journal of Ecology, 2013, 101(6):1520-1530.
[5] Augspurger C K. Phenology, flowering synchrony and fruit set of six Neotropical shrubs[J].Biotropica, 1983, 15(4):257-267.
[6] Pickering C M. Variation in flowering parameters within and among five species of Australian alpine Ranunculus[J].Australian Journal of Botany, 1995, 43(1): 103-112.
[7] Primack R B. Relationships among flowers, fruits and seeds[J]. Annual Review of Ecology & Systematics, 1987, 18(1): 409-430.
[8] 刘志民, 闫巧玲, 马君玲, 等. 生殖物候与草甸草地多年生植物的消长[J].生态学报, 2006,26(3):773-779.
[9] Fenner M. The phenology of growth and reproduction in plants[J]. Perspectives in Plant Ecology and Systematics, 1998, 1(1):78-91.
[10] 陈敦宪. 谈果实各组成部分的基因型和表现型[J].生物学教学,1999(12):19-20.
[11] 毛祖美. 新疆植物志:第二卷 第一分册 [M]. 乌鲁木齐:新疆科技卫生出版社, 1994.
[12] 蔡丹红. 两种藜科植物的生殖物候及其种子萌发行为[D].乌鲁木齐:新疆农业大学, 2015.
[13] 布海丽且姆·阿卜杜热合曼, 严成, 魏岩. 小蓬的开花生物学及果实形态发育特性[J]. 新疆农业大学学报, 2012, 35(4):306-311.
[14] 张艳敏, 魏岩, 严成. 亚麻叶碱蓬(Suaeda linifolia)的物候及结实格局[J]. 新疆农业大学学报, 2010, 33(3):187-191.
[15] 郭红超. 木地肤的生殖生物学研究[D]. 乌鲁木齐:新疆农业大学, 2014.
[16] 马文宝, 施翔, 张道远, 等. 准噶尔无叶豆的开花物候与生殖特征[J]. 植物生态学报, 2008, 32(4):760-767.
[17] 杨旭. 荒漠植物骆驼蓬繁殖生物學的研究[D]. 石河子:石河子大学, 2018.
[18] Philbrick C T, Retana A N. Flowering phenology, pollen flow and seed production in Marathrum rubrum(Podostemaceae)[J]. Aquatic Botany, 1998, 62(3): 199-206.
[19] Lloyd D G, Webb C. The avoidance of interference between the presentation of pollen and stigmas in angiosperms Ⅰ. Dichogamy[J]. New Zealand Journal of Botany, 1986, 24(1):135-162.
[20] Harder L D, Barrett S C H. Pollen dispersal and mating patterns in animal-pollinated plants[M]. New York: Chapman and Hall, 1996:140-190.
[21] Evanhoe L, Galloway L F. Floral longevity in Campanula americana(Campanulaceae): a comparison of morphological and functional gender phases[J]. American Journal of Botany, 2002, 89(4):587-591.
[22] 高蕊, 魏岩. 荒漠植物角果藜的地上地下结果性[J]. 云南植物研究, 2007(3): 300-302.
