荣成近岸养殖海域浮游植物群落结构及与环境因子的关系*
2019-09-27潘玉龙沙婧婧徐东会王泰森刘旭东
潘玉龙 刘 潇 沙婧婧 徐东会 王泰森 刘旭东
荣成近岸养殖海域浮游植物群落结构及与环境因子的关系*
潘玉龙 刘 潇 沙婧婧①徐东会 王泰森 刘旭东
(1. 国家海洋局北海环境监测中心 国家海洋局海洋溢油鉴别与损害评估技术重点实验室 山东省海洋生态环境与防灾减灾重点实验室 青岛 266033)
基于2016年4个季节的生态调查数据,研究了山东荣成近岸养殖海域浮游植物群落结构,同时,应用典范对应分析(CCA)讨论了环境因子对荣成近岸养殖海域浮游植物群落结构的影响。4个季度共发现浮游植物66种,分隶于硅藻、甲藻及金藻3个植物门、29个属;浮游植物优势种包括中肋骨条藻()、具槽直链藻()、日本星杆藻()、扭链角毛藻()、尖刺菱形藻()及劳氏角毛藻(),具有明显的季节演替现象。春季日本星杆藻在荣成湾大量出现使其成为优势种。4个季节浮游植物丰度变化范围为(16.9~251.7)× 104cells/m3,平均为119.3×104cells/m3,以夏季最高,秋季最低;全年浮游植物多样性指数(¢)、均匀度指数()、丰富度指数()和优势度指数(D)的变化范围分别为1.60~2.13、0.40~0.60、0.58~0.79和0.70~ 0.84,多样性指数以夏季最高,冬季最低,但均处在较好水平;CCA分析结果表明,海水溶解氧(DO)、硅酸盐(SiO32–-Si)、铵盐(NH+4)、温度和pH等环境因子为影响该海域浮游植物群落结构的主要因子,除了季节影响因素以外,当地多元化的养殖模式可能是导致目前浮游植物群落结构的重要因素。
养殖海域;浮游植物;群落结构;环境因子;典范对应分析
荣成市位于山东半岛最东端,三面环海,其附近海域是我国北方最主要的海水养殖基地之一,荣成湾养殖区以及桑沟湾–镆铘岛养殖区等均为山东省重点养殖海区。统计结果显示,2016年荣成市海水养殖面积和养殖产量分别占威海市总海水养殖面积和养殖产量的49.6%和41.8%。
海水养殖对周围海洋生态环境的影响是非常复杂的,其很大程度上取决于养殖种类、养殖模式、养殖密度、饵料类型及养殖区水文条件等。但大规模的海水养殖活动会造成海水富营养化、海水溶解氧(DO)含量降低以及浮游植物群落结构发生变化,浮游植物大量增殖引发赤潮,反过来又会对养殖活动产生灾难性的破坏。关于桑沟湾海域的浮游植物群落结构特征的研究已有不少(刘慧等, 2003; 李超伦等, 2010; 钱罡等, 2017),荣成月湖(帅莉等, 2003)和荣成俚岛人工鱼礁区(刘长东等, 2016)附近海域的浮游植物群落结构也有报道,但未见北部荣成湾养殖区海域浮游植物群落结构的相关研究,更少见有关整个荣成附近养殖海域浮游植物群落结构的调查、附近养殖海域浮游植物群落结构与环境因子关系的分析研究。荣成湾与桑沟湾养殖地位与养殖模式相似,但地形特征和营养盐结构却与桑沟湾差异较大(谢琳萍等, 2013),2个海湾共同作用下可能会导致整个荣成近岸养殖海域浮游植物群落结构出现新的格局。本研究基于2016年2、5、8、11月4个季节对整个荣成海域的水生生态综合调查,全面分析整个荣成海域浮游植物群落结构的季节变化特征,并通过典范对应分析(CCA)进一步研究该海域浮游植物群落结构与环境因子的关系,分析不同季节浮游植物与环境因子和海水养殖的关系,为阐明海水养殖对不同海湾近海生态系统的影响,建立科学合理的生态养殖模式提供基本依据。
1 材料与方法
1.1 站位布设与样品采集
于2016年2、5、8和11月对山东荣成附近海域进行4个季节的大面积调查。共设10个调查站位(图1),浮游植物取样按照《海洋监测规范》(GB/ T17378.7–2007)进行,采用浅水Ⅲ型浮游生物网,于各站自水体底层至表层垂直拖取。样品采用5%福尔马林溶液固定保存,带回实验室后在倒置显微镜下鉴定和计数。
测定的环境参数包括海水表层温度(SST)、透明度(TRA)、表层盐度(S)、酸度(pH)、溶解氧(DO)、化学需氧量(COD)、磷酸盐(PO43–-P)、亚硝酸盐(NO–2-N)、硝酸盐(NO–3-N)、铵盐(NH+4)、硅酸盐(SiO32–-Si)、总磷(TP)、总碱度(ALK)和总氮(TN)。水质样品的采集和参数测定方法参照国标《海洋监测规范》(GB/T17378.4–2007)进行。
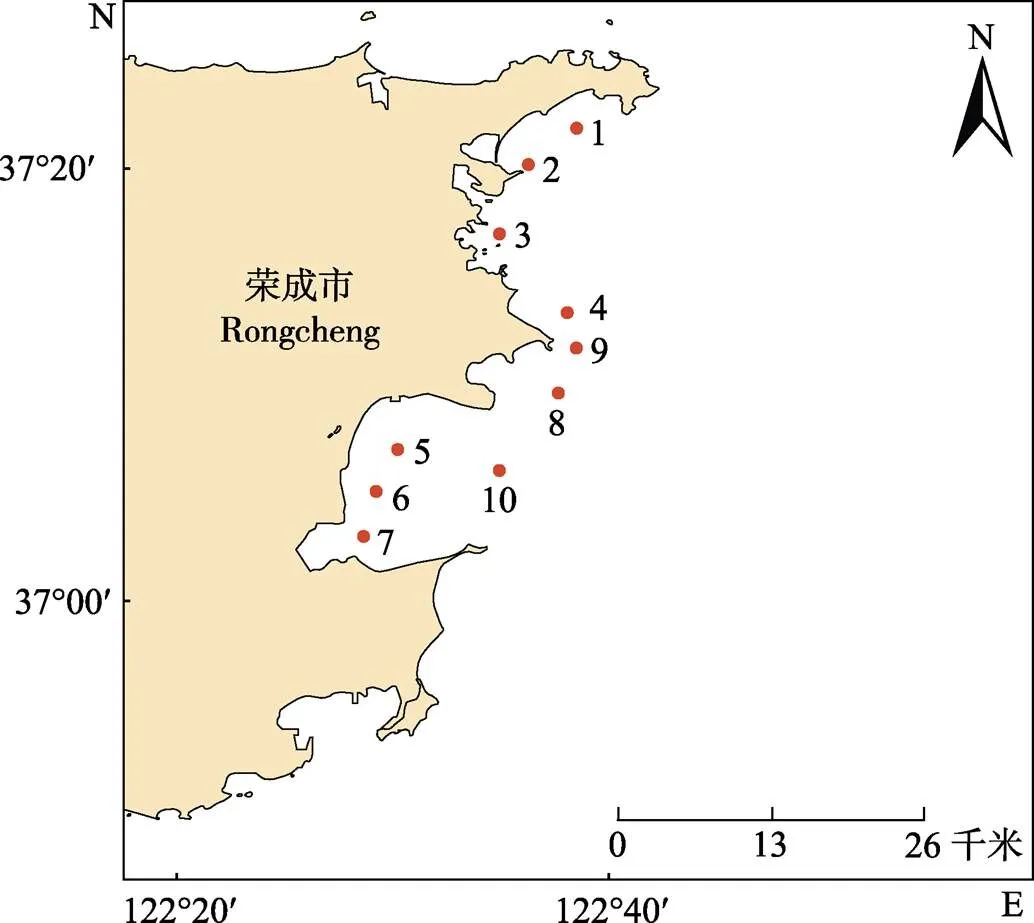
图1 荣成附近养殖海域调查站位
1.2 群落结构特征指数
浮游植物群落结构多样性指数(¢)、均匀度指数()、丰富度指数()和优势度指数(2)的计算公式如下:
Shannon-Wiener多样性指数():
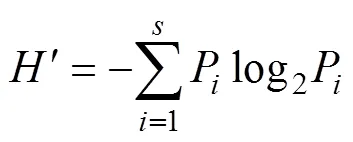
Pielou均匀度指数():=/log2
丰富度指数():=(-1)/log2
优势度指数(2):2=(1+2)/NT
式中,P=n/(n为第个物种的个体数,为全部物种的个数);为浮游植物种类数;1为样品中第1优势种的个体数;2为样品中第2优势种的个体数;NT为样品中的总个体数。
浮游植物优势种根据优势度计算结果判定,计算公式:
=P×f
式中,P=n/(n为第个物种的个体数,为全部物种的个数);f为第个物种在各站位出现的频率,定义优势度大于0.02的物种为优势种(陈亚瞿等, 1995)。
1.3 典范对应分析(CCA)
CCA是一种非线性多元统计分析方法,适用于研究一个变量与多个因子的关系,是环境生态学研究的重要方法(Ter Braak, 1986)。本研究采用Canoco for Windows 4.5软件对浮游植物密度和环境因子进行典范对应分析,根据以下2个原则筛选物种:(1)物种的站位出现率大于15%;(2)物种优势度大于0.001。共选取了18种浮游植物进行CCA分析,物种矩阵经log(+1)转换。
2 结果
2.1 环境数据分析
调查海域海水水质总体良好,无机氮含量在不同季节均存在超一类《海水水质标准》(GB 3097–1997)的现象,主要是由于NO–3-N含量较高所致,其他环境参数均符合一类《海水水质标准》。各参数统计结果见表1。
2.2 浮游植物群落结构
2.2.1 种类组成和优势种 调查海域4个季节共发现浮游植物66种,隶属于硅藻门(Diatom)、甲藻门(Pyrrophyta)及金藻门(Chrysophyta)3个植物门、29个属,生态类型主要为温带近岸种。其中,硅藻58种,占浮游植物总种数的87.9%,以角毛藻属()物种种类最多,为14种,占总种数的10.6%;甲藻为7种;金藻为1种,占1.5%。浮游植物种名录见表2。
表1 荣成养殖区海域各环境参数统计结果
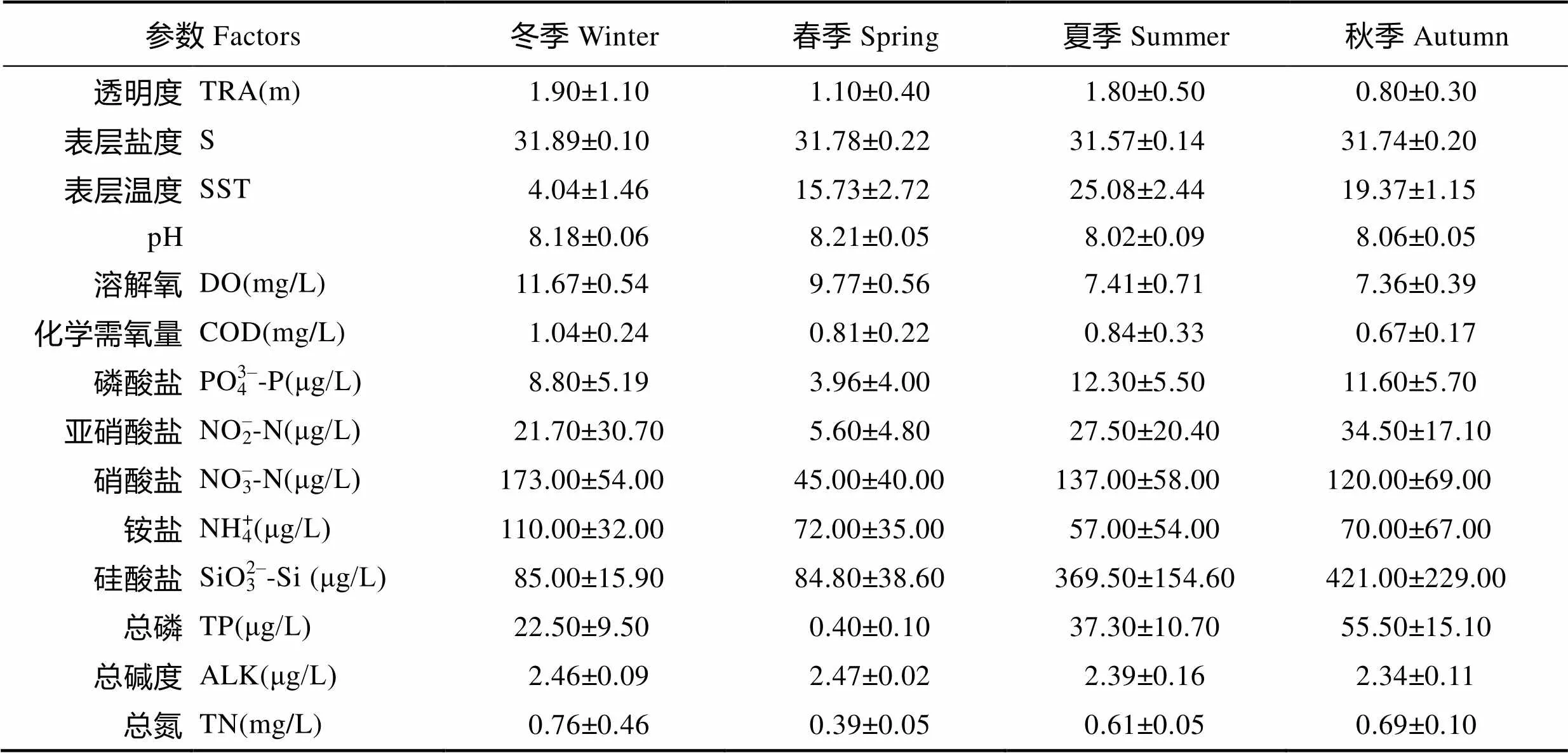
Tab.1 The results of environmental parameters in Rongcheng adjacent waters for culture
调查海域浮游植物优势种4个季节均为硅藻。在夏季,扭链角毛藻()和旋链角毛藻()的大量出现,使得该海域浮游植物优势种发生了变化,其他3个季节具槽直链藻()均为优势种之一,冬季和春季中肋骨条藻()也是浮游植物优势种,春季日本星杆藻()在荣成湾大量出现,使其成为该季节浮游植物优势种(表3),这与李超伦等(2010)、钱罡等(2017)调查结果有所不同。
2.2.2 丰度 从表4可以看出,荣成养殖海域4个季节浮游植物平均丰度变化范围为(16.9~251.7)× 104cells/m3,平均丰度为119.3×104cells/m3,表现出明显的季节变化,其中,夏季(8月)最高,冬季(3月)次之,秋季(11月)最低,这与慕建东等(2009)的研究结果相一致。
2.2.3 多样性指数 4个季节的浮游植物多样性指数平均值见表5。从表5可以看出,该海域全年浮游植物多样性指数变化范围为1.60~2.13,根据生物多样性阈值评价标准(陈清潮等, 1994),调查海域全年浮游植物生物多样性均处在较好水平。
多样性指数最高出现在夏季,同时,夏季的均匀度指数最高,丰度和优势度指数最低,表明夏季浮游植物多样性较高,相对而言,冬季浮游植物多样性较低。
2.2.4 浮游植物的典范对应分析(CCA) 经筛选,用于CCA排序的浮游植物种类包括18种(表6),环境因子14项,具体包括海水SST、TRA、表层盐度、pH、DO、COD、PO43–-P、NO–2-N、SiO32–-Si、NH+4、NO–3-N、TP、TLK以及TN。
CCA二维排序图中(图2),前2个排序轴的特征值为0.907和0.795,物种排序轴和环境因子轴之间的相关系数分别为0.991和0.975,物种变异累积分别为25.6%和48.0%,物种环境变异累积分别为29.8%和55.8%,说明前2个排序轴能在很大程度上反映浮游植物种类与环境因子间的关系。在CCA排序中,与第1排序轴呈最大正相关的因子为DO,相关系数为0.890,其次为NH+4和pH,其相关系数分别为0.863和0.853;SiO32–-Si与第1排序轴呈最大负相关,相关系数为–0.940,其次为海水SST和TP,其相关系数分别为–0.739和–0.737;NO–2-N、SiO32–-Si、ALK等环境因子也与第1排序轴呈较大的负相关关系。SST与第2排序轴呈最大正相关,相关系数为0.583;ALK、SiO32–-Si和SST均与第2排序轴呈一定的正相关关系;其余环境因子均与第2排序轴呈负相关关系,其中,TP呈最大负相关,其相关系数为–5.454;其次为TN,其相关系数为–4.602。
表2 调查海域浮游植物种名录

Tab.2 List of phytoplankton species in the surveyed area
从图2可以看出,仅有日本星杆藻分布在图2的右上方。从第1排序轴上看,该物种与海水DO、NH+4和pH呈较大的正相关;从第2排序轴上看,与SST呈较大的正相关。
在前2个轴中,NO–2-N、PO43–-P、TP、COD和NO–3-N均与日本星杆藻呈负相关关系,而这些环境因子均与位于图2左下角的劳氏角毛藻呈正相关关系。
中肋骨条藻、具槽直链藻、圆筛藻、棱角海链藻、卡氏角毛藻、柔弱根管藻、加氏星杆藻、诺氏海链藻和刚毛根管藻均位于图2的右下方,分布较集中,说明它们与DO、NH+4、pH、TRA和TN含量呈正相关,倾向于生长在这些因子较高的区域,但与温度、ALK和SiO32–-Si呈负相关关系。旋链角毛藻、布氏双尾藻、角毛藻、扭链角毛藻、尖刺菱形藻和丹麦细柱藻较密集分布于图2的左上方,这些物种的分布与海水温度、SiO32–-Si含量及ALK呈正相关关系。
表3 荣成养殖区海域不同季节的浮游植物优势种
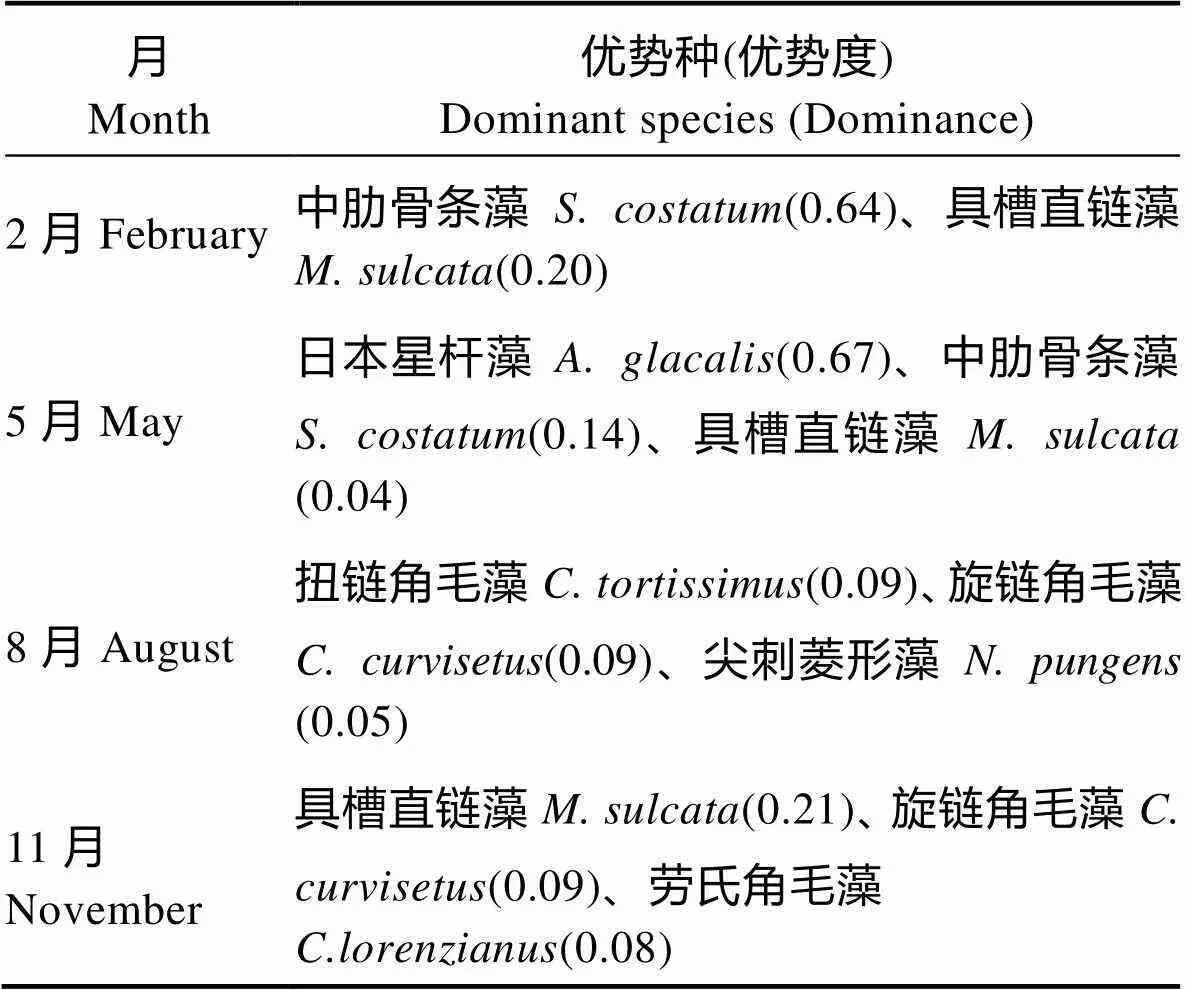
Tab.3 Dominant species of phytoplankton in different seasons of the surveyed area
表4 荣成养殖区海域浮游植物丰度结果
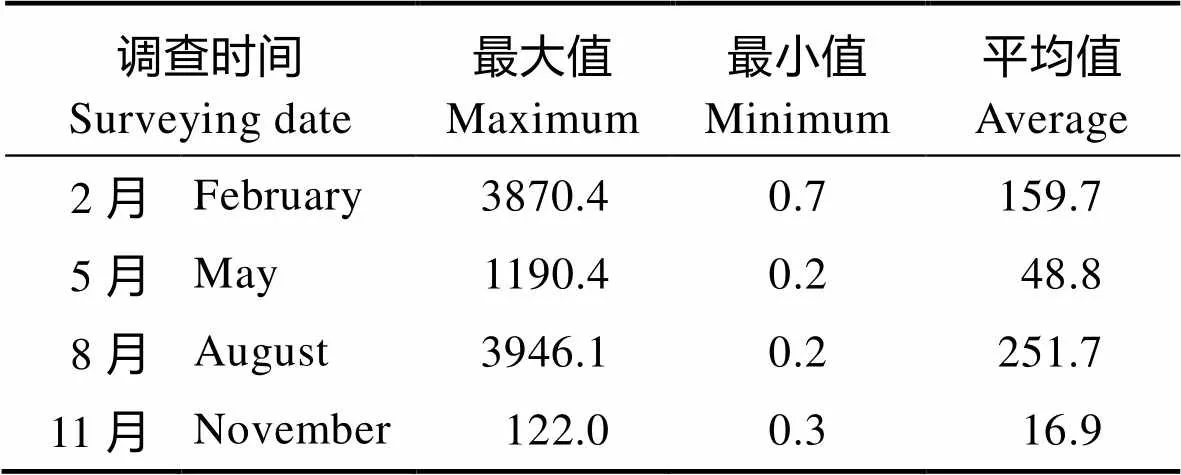
Tab.4 The abundance of phytoplankton in Rongcheng adjacent waters for culture (×104 cells/m3)
表5 调查海域不同季节的浮游植物群落结构特征指数
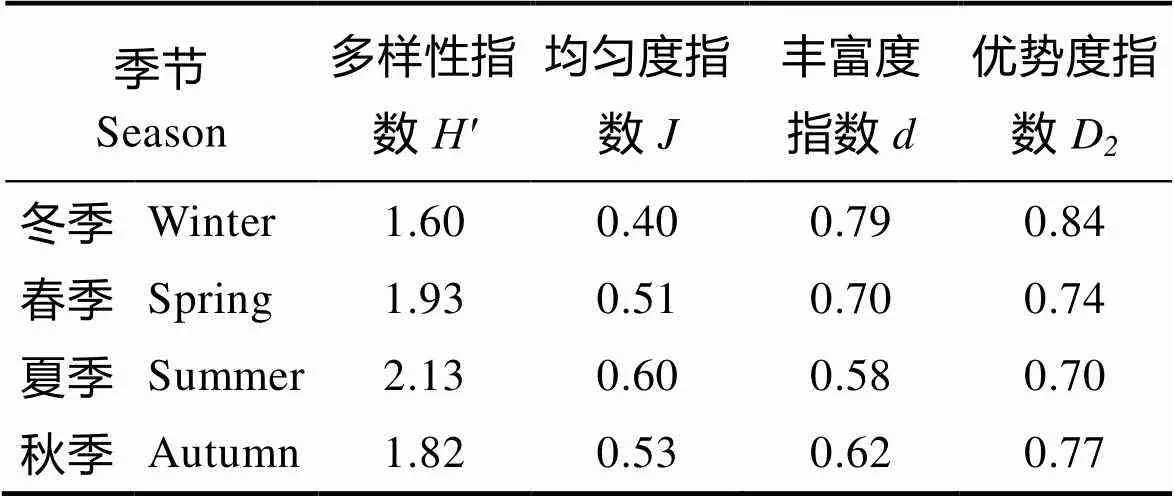
Tab.5 Phytoplankton diversity imdices of different seasons in thesurveyed area
表6 CCA排序中浮游植物种类编号

Tab.6 The code of phytoplankton in CCA
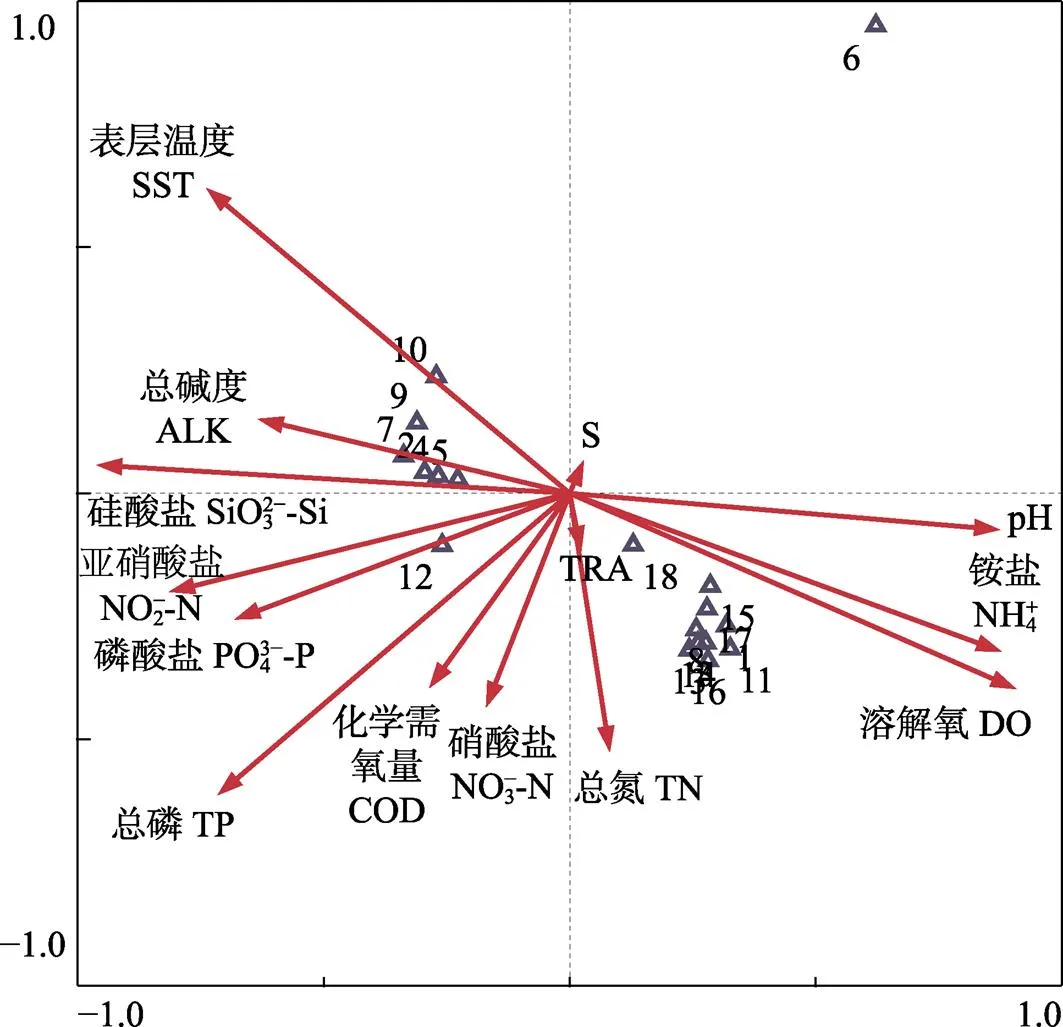
图2 主要浮游植物种类与环境因子的CCA排序
3 讨论
3.1 浮游植物群落结构
整个荣成养殖区附近海域浮游植物种类组成与桑沟湾养殖区海域一致,以近岸广布种为主,硅藻占绝对优势,优势种存在季节演替现象,这与李超伦等(2010)的研究结果相同。钱罡等(2015年)研究了桑沟湾海域春季浮游植物的群落结构,浮游植物优势种为具槽直链藻、离心列海链藻和布氏双尾藻。本研究中,春季北部荣成湾内日本星杆藻大量出现,使其成为整个调查海域的绝对优势种,但具槽直链藻仍然保持了较高的密度,成为优势种类之一;本次调查浮游植物密度平均值低于2015年春季调查结果,但与李超伦等(2010)和慕建东等(2009)的研究结果较接近,可能是由于2015年采用的浮游植物调查方法不同所致。
本次调查浮游植物多样性指数与桑沟湾的历史研究相比较也在正常波动范围之内(宋洪军等, 2007; 慕建东等, 2009)。以上研究表明,荣成养殖区附近海域浮游植物群落结构与桑沟湾总体基本保持一致,仅春季优势种不同于历史调查结果,表明近些年荣成养殖区附近海域浮游植物群落结构基本稳定。
3.2 浮游植物群落结构与环境因子的关系
影响浮游植物生长繁殖的环境因子比较多,机制也比较复杂(孙军等, 2005; 杨阳等, 2016)。本研究结果显示,荣成近岸养殖海域浮游植物群落结构主要受DO、SiO32–-Si、NH+4、温度和pH等因素影响。其中,DO、NH+4、pH等促进优势种中肋骨条藻、具槽直链藻以及日本星杆藻的生长,而温度和SiO32–-Si含量则与这些种类的密度成负相关;夏季出现的优势种类(角毛藻类)则与海水温度、SiO32–-Si含量呈正相关关系。
3.3 浮游植物群落结构与海水养殖的关系
调查海域的养殖模式每年相对固定,附近海域主要为海带()和贝类养殖区域,其中,贝类养殖主要在近岸水较浅的湾内进行,海带养殖则集中在外侧水深流急的海域,养殖方式均为筏式养殖,贝类养殖起始于8月,一般至翌年的5月全部结束(傅明珠等, 2013),而海带养殖时间一般为每年11月至翌年5月,5月海带开始收获,夏季来临之前收获全部完成(史洁等, 2010)。在冬季,海带幼苗相对较小,海水中DO和营养盐含量相对较高,浮游植物大量繁殖,因此,冬季浮游植物丰度较高;4~5月为海带临收获季节,海带生物量和生物密度达到最大,海水中的DO含量相对较高,而营养盐含量较低,王玉珏等(2008)研究表明,日本星杆藻可以在低氮条件下表现出竞争优势,迅速分裂生长,日本星杆藻为绝对优势种;夏季海带收割后,营养盐消耗骤减,补充明显,另外,残留的部分藻体腐败变质,也释放出大量的营养盐(张继红等, 2010),因此,浮游植物营养盐限制压力被缓解,开始大量生长繁殖(徐东会等, 2017),且海水温度较高,浮游植物优势种演替为角毛藻类;之后随着营养盐的消耗,海水温度的降低,秋季浮游植物密度下降,优势种又成为具槽直链藻。
关于贝类养殖对浮游植物群落结构影响的研究较多(王俊等, 2001; 董婧等, 2003),但研究结果并不相同,荣成近岸养殖海域贝类养殖高峰期为夏季,此时,海带已经收割完毕,这期间,贝类排泄物以及沉积物成为海水营养盐的重要来源(孙珊等, 2010),海水pH和DO含量也因为高密度养殖贝类的活动而降低(陶平等, 2005),而SiO32–-Si含量较高,这导致夏季角毛藻类大量增殖,另外,养殖贝类的快速生长消耗了大量浮游动物幼体,从而间接促进了浮游植物的增殖(刘萍等, 2013)。因此,夏季高密度的贝类养殖可能促进了浮游植物的生长繁殖,这与史洁等(2010)的研究结果相一致。
Wu RSS. The environmental impact of marine fish culture: Towards a sustainable future. Marine Pollution Bulletin, 1995, 1(31): 95–166
Chen YQ, Xu ZL, Wang YL,. An ecological study on zooplankton in plume front zone of Changjiang(Yangzi) River Estuarine area: I. Biomass distribution of dominant species. Journal of Fishery Science of China, 1995, 2(1): 49–58 [陈亚瞿, 徐兆礼, 王云龙, 等. 长江口河口锋区浮游动物生态研究: I. 生物量及优势种的平面分布. 中国水产科学, 1995, 2(1): 49–58]
Chen QC, Huang LM, Yin JQ. Research of diversity of zooplankton in Nansha Islands areas. Beijing: China Ocean Press, 1994 [陈清潮, 黄良民, 尹建强. 南沙群岛海区浮游动物多样性研究. 北京: 海洋出版社, 1994]
Dong J, Bi YP, Wang WB,. Effect of high density bivalve rafts cultivation on phytoplankton community. Marina Fisheries Research, 2003, 24(3): 50–54 [董婧, 毕远溥, 王文波, 等. 小窑湾高密度贝类筏式养殖对浮游植物群落的影响. 海洋水产研究, 2003, 24(3): 50–54]
Fu MZ, Pu XM, Wang ZL,. Integrated assessment of mariculture ecosystem health in Sanggou Bay. Acta Ecologica Sinica, 2013, 33(1): 238–248 [傅明珠, 蒲新明, 王宗灵, 等. 桑沟湾养殖生态系统评价. 生态学报, 2013, 33(1): 238–248]
Li CL, Zhang YS, Sun S,. Phytoplankton species composition, quantity distribution and seasonal variation in the Sanggou Bay. Progress in Fishery Sciences, 2010, 31 (4): 1–8 [李超伦, 张永山, 孙松, 等. 桑沟湾浮游植物种类组成、数量分布及其季节变化. 渔业科学进展, 2010, 31(4): 1–8]
Liu CD, Yi J, Guo XF,. Phytoplankton community structure in artificial reef area around Lidao of Rongcheng, and its relationship with environmental factors. Periodical of Ocean University of China(Natural Science), 2016, 40(3): 50–59 [刘长东, 易坚, 郭晓峰, 等. 荣成俚岛人工鱼礁区浮游植物群落结构及其与环境因子的关系. 中国海洋大学学报(自然科学版), 2016, 40(3): 50–59]
Liu H, Fang JG, Dong SL,. Study on phytoplankton in raft-culture areas of Laizhou Bay and Sanggou Bay . Marine Fisheries Research, 2003, 24(2): 9–17 [刘慧, 方建光, 董双林, 等. 莱州湾和桑沟湾养殖海区浮游植物的研究. 海洋水产研究, 2003, 24(2): 9–17]
Liu P, Song HJ, Fu MZ,. Seasonal variability of zooplankton community characteristics in the Rongcheng Bay contiguous waters. Acta Oceanologica Sinica, 2013, 35(4): 168–175 [刘萍, 宋洪军, 傅明珠, 等. 荣成湾毗邻海域浮游动物群落季节分布特征. 海洋学报, 2013, 35(4): 168–175]
Mu JD, Dong W, Chen BJ,. Progress of ecological characteristics of phytoplankton in Sanggou Bay. Progress in Fishery Sciences, 2009, 30(3): 54–58 [慕建东, 董玮, 陈碧鹃, 等. 桑沟湾浮游植物生态特征. 渔业科学进展, 2009, 30(3): 91-96]
Qiang G, Wei YQ, Sun J. Study on phytoplankton communities in the Sanggou Bay in spring, 2015. Marine Sciences, 2017, 41(2): 44–52 [钱罡, 魏玉秋, 孙军. 2015年春季桑沟湾浮游植物群落研究. 海洋科学, 2017, 41(2): 44–52]
Qiang SB, Chen GW, Tang TY. Study on the phytoplankton from the offshore waters of Rongcheng, Shandong. Journal of Shandong College of Oceanology (Natural Science), 1981, 11(3): 52–70 [钱树本, 陈国蔚, 汤庭耀. 山东省荣城县近海浮游植物的研究. 山东海洋大学学报(自然科学版), 1981, 11(3): 52–70]
Shi J, Wei H, Zhao L,. Study on ecosystum model for multi–species culture in Sanggou Bay: I. Establishment of the culture ecosystum model and sensitivity analyses of parameters. Progress in Fishery Sciences, 2010, 31(4): 26–35 [史洁, 魏皓, 赵亮, 等. 桑沟湾多元养殖生态模型研究: I. 养殖生态模型的建立和参数敏感性分析. 渔业科学进展, 2010, 31(4): 26–35]
Shi J, Wei H, Zhao L,. Study on ecological model of multiple breeding in Sanggou Bay.Ⅱ. Progress, ecological environment simulation and nutrients cycling. Progress in Fishery Sciences, 2010, 31(4): 36–42 [史洁, 魏皓, 赵亮, 等. 桑沟湾多元养殖生态模型研究:Ⅱ.生态环境模拟与生源要素循环. 渔业科学进展, 2010,31(4): 36–42]
Shuai L, Yin X, Yang XN,. Study on the phytoplankton and zooplankton in the Yuhu Inlet, Shandong Peninsula. Journal of Qingdao University, 2003, 18(4): 70–75 [帅莉, 殷效, 杨小妮, 等. 山东荣成月湖浮游动植物的研究.青岛大学学报, 2003, 18(4): 70–75]
Song HJ, Li RX, Wang ZL,. The interannual variation of phytoplankton diversity in Sanggou Bay. Advances in Marine Science, 2007, 25(3): 332–339 [宋洪军, 李瑞香, 王宗灵, 等. 桑沟湾浮游植物多样性年际变化. 海洋科学进展, 2007, 25(3): 342–346]
Sun S, Liu SM, Ren JL,. Distribution features of nutrients and flux across the sediment—water interface in the Sanggou Bay. Acta Oceanologica Sinica, 2010, 32(6): 108–117 [孙珊, 刘素美, 任景玲, 等. 桑沟湾养殖海域营养盐和沉积物. 水界面扩散通量研究. 海洋学报, 2010, 32(6): 108–117]
Sun J, Liu DY. Net-phytoplankton community of the Bohai Sea in the autumn of 2000. Acta Oceanologica Sinica, 2005, 27(3): 124–132 [孙军, 刘东艳. 2000年秋季渤海的网采浮游植物群落. 海洋学报, 2005, 27(3): 124–132]
Tao P, Ding YS, Li BT,. Study on the effects of shellfish increasing on aquatic environment by laboratory simulation. Acta Scientiae Circumstantiae, 2005, 25(6): 829–835 [陶平, 丁永生, 李柏弢, 等. 增加投放量对贝类养殖水环境影响的模拟研究. 环境化学, 2005, 25(6): 829–835]
Ter Braak CJF. Canonical correspondence analysis: A new eigenvector method for multivariate direct analysis. Ecology, 1986, 67(5): 1167–1179
Wang J, Jiang ZH, Dong SL. Role of filter-feeding bivalves in proliferation of phytoplankton community. Chinese Journal of Applied Ecology, 2001, 12(5): 765–768 [王俊, 姜祖辉, 董双林. 滤食性贝类对浮游植物群落增殖作用的研究. 应用生态学报, 2001, 12(5): 765–768]
Wang YJ, Hong HS, Wang DZ,. Response of phytoplankton to nutrients addition in the upwelling regions of the Taiwan Strait. Acta Ecologica Sinica, 2008, 28(3): 1321—1327 [王玉珏, 洪华生, 王大志, 等. 台湾海峡上升流区浮游植物对营养盐添加的响应. 生态学报, 2008, 28(3): 1321—1327]
Wu WG, Zhang JH, Wang W,. Variation of Chl-concentration and its control factors pre-and-post-the-harvest of kelp in Sanggou Bay. Journal of Fisheries of China, 2015, 39(8): 1178–1186 [吴文广, 张继红, 王巍, 等. 桑沟湾水域叶绿素在海带收获前后的变化及其影响因素. 水产学报, 2015, 39(8): 1178–1186]
Xie LP, Pu XM, Sun X,. Analysis on the temporal and spatial distribution of nutrients and the influence factors in Rongcheng Bay. Marine Science Bulletin, 2013, 32(1): 20–27 [谢琳萍, 蒲新明, 孙霞, 等. 荣成湾营养盐的时空分布特征及其影响因素分析. 海洋通报, 2013, 32(1): 20–27]
Xu DH, Chen BJ, Jiang T,. Temporal and spatial variation characteristics of water nutrients and eutrophication evaluation in aquaculture water areas of Sanggou Bay. Progress in Fishery Sciences, 2017, 38(3): 1–11 [徐东会, 陈碧娟, 江涛, 等. 桑沟湾养殖海域营养盐时空分布特征及富营养化评价. 渔业科学进展, 2017, 38(3): 1–11]
Yang Y, Sun J, Guang XY,. Seasonal variation of netz- phytoplankton community in Bohai Sea. Marine Science Bulletin, 2016, 35(2): 121–131 [杨阳, 孙军, 关翔宇, 等. 渤海网采浮游植物群集的季节变化. 海洋通报, 2016, 35(2): 121–131]
Zhang JH, Jiang ZJ, Wang W,. Seasonal distribution and variation of nutrients and nutrients limitation in Sanggou Bay. Progress in Fishery Sciences, 2010, 3l(4): 16–25 [张继红, 蒋增杰, 王巍, 等. 桑沟湾营养盐时空分布及营养盐限制分析. 渔业科学进展, 2010, 3l(4): 16–25]
Influence of Environmental Factors on Phytoplankton Community Structure and Its Relationship with Coastal Aquaculture in the Waters Adjacent to Rongcheng
PAN Yulong, LIU Xiao, SHA Jingjing①, XU Donghui, WANG Taisen, LIU Xudong
(1. North China Sea Environmental Monitoring Center, State Oceanic Administration; Laboratory of Marine Spill Oil Identification and Damage Assessment Technology, State Oceanic Administration, Shandong Provincial Key Laboratory of Marine Ecological Environment and Disaster Prevention and Mitigation, Qingdao 266033)
The phytoplankton community structure in the coastal aquaculture area of Rongcheng was analyzed based on the results of four surveys during 2016. The influence of environmental factors on the phytoplankton community in the Rongcheng coastal aquaculture area was discussed using canonical correlation analysis. The results identified a total of 66 species that belong to 29 genera of 3 phyla. The dominant species of phytoplankton include,,,,,andwith seasonal succession. In the spring.increased quickly in Rongcheng Bay to make itthe most important dominant species of this season, but not in Sanggou Bay, when compared with the results of previous studies. The abundance of phytoplankton varied from 16.9×104to 251.7×104cells/m3over the 4 seasons, the average density of whole year was 119.3×104cells/m3, which was the highest in summer and the lowest in autumn. The Shannon-Wiener diversity index and Pielou’s index range from 1.60 to 2.13 and 0.40 to 0.60, respectively, and the diversity of phytoplankton was the highest in the summer, and the mode of the phytoplankton structure changed with the seasons, but the levels of phytoplankton remained normal all year round. Canonical correlation analysis showed that the most important factors affecting phytoplankton community structure are dissolved oxygen, silicate, NH+4, sea surface temperature and pH. These may promote or inhibit the growth of specific species and cause the rise of dominant species. Large-scale kelp farming in the area changed the amount of nutritional salt, high density shellfish farming consumed the natural enemies of phytoplankton, and shellfishexcretion also changed the composition of nutritional salts in the coastal waters; therefore, local aquaculture in the Rongcheng coastal waters may be an important factor in the pattern of phytoplankton community structure in addition to the influences of seasonal changes.
Aquaculture waters; Phytoplankton; Community structure; Environmental factors; Vanonical correlation analysis
SHA Jingjing, E-mail: panyulong@bhfj.gov.cn
* 国家重点研发计划海洋环境安全保障重点专项(2016YFC1402305)、国家海洋局海洋溢油鉴别与损害评估技术重点实验室开放基金(201701)、山东省海洋生态环境与防灾减灾重点实验室开放基金(201602)和国家海洋局海洋生态环境科学与工程重点实验室开发基金(MESE–2018–02)共同资助[This work was supported by the Joint Fund Project of National Key Research and Development Program Marine Environment Security Guarantee Key Special Project (2016YFC1402305), Open Fund Project of Laboratory of Marine Spill Oil Identification and Damage Assessment Technology (201701), Open Fund Project of Shandong Provincial Key Laboratory of Marine Ecological Environment and Disaster Prevention and Mitigation (201602), and Open Fund Project of State Oceanic Administration (MESE–2018–02)]. 潘玉龙, E-mail: ylpan2006@163.com
沙婧婧,E-mail: panyulong@bhfj.gov.cn
2018-07-10,
2018-08-17
Q948
A
2095-9869(2019)05-0026-08
10.19663/j.issn2095-9869.20180710001
http://www.yykxjz.cn/
潘玉龙, 刘潇, 沙婧婧, 徐东会, 王泰森, 刘旭东. 荣成近岸养殖海域浮游植物群落结构及与环境因子的关系. 渔业科学进展, 2019, 40(5): 26–33
Pan YL, Liu X, Sha JJ, Xu DH, Wang TS, Liu XD. Influence of environmental factors on phytoplankton community structure and its relationship with coastal aquaculture in the waters adjacent to Rongcheng. Progress in Fishery Sciences, 2019, 40(5): 26–33
(编辑 陈 严)
