科尔沁沙地不同苜蓿品种越冬期根颈耐寒生理机制的研究
2019-09-24郝培彤宁亚明高秋刘芳李玉龙张尚雄郑桂亮王显国
郝培彤,宁亚明,高秋,刘芳,李玉龙,张尚雄,郑桂亮,王显国*
(1.中国农业大学草业科学系,北京 100193;2.全国畜牧总站,北京 100125;3.天津市饲草饲料工作站,天津 300210)
紫花苜蓿(Medicagosativa) 被誉为“牧草之王”,具有生物固氮能力,其根系十分发达可减少地表径流,具很强的水土保持和防风固沙能力,《全国苜蓿产业发展规划 (2016-2020年)》提出到 2020 年全国优质苜蓿产量达到5.4×106t[1]。科尔沁沙地东起吉林省双辽县,西至内蒙古赤峰市翁牛特旗,南北介于燕山北部黄土丘陵和大兴安岭东麓丘陵,地跨内蒙古、吉林、辽宁3个省区,总面积大约5.17×104km2,为我国北方重要的农牧业生产基地。近年来,随着以奶业为龙头的草食家畜业迅猛发展,依托较好的自然条件、政策与区位优势,本区苜蓿生产得到了快速的发展,现已成为我国机械化程度最高、集中连片生产面积最大的新兴苜蓿优势产区,其中核心产地-赤峰市阿鲁科尔沁旗现有具备喷灌设施的紫花苜蓿生产基地5×104hm2。
紫花苜蓿作为多年生牧草,越冬能力是影响其翌年产量的重要因素,阿旗地区昼夜温差大,以2月为例 (近7年数据),白天温度可达10 ℃、夜间-25 ℃,土壤不断地发生冻融交替,加之该地区冬季干旱降水少极易导致地表附近的根颈脱水死亡丧失再生能力。冬季干旱造成的苜蓿非正常死亡主要表现为植株根颈部位失水收缩,较严重的根颈出现黑斑坏死甚至整个根部呈干枯状,一旦根颈受冻产生黑斑甚至变黑、腐烂时便不会返青,对越冬率及翌年的生长产生影响[2]。
秋眠级可以作为判断苜蓿抗寒性的依据,不同秋眠级苜蓿根颈特性差异显著[3]。王龙昌等[4]研究指出不同苜蓿品种越冬期间根颈直径、枝条再生数及越冬后产量都与土壤含水量呈正相关。方强恩[5]研究表明紫花苜蓿根颈芽细胞在越冬期间会发生一系列适应低温的变化提高根颈芽的抗寒能力。研究表明抗寒锻炼可显著提高紫花苜蓿的抗寒性[6],秋眠性紫花苜蓿品种Evolution经抗寒锻炼后可显著提高根颈可溶性糖含量,其中蔗糖、棉子糖、水苏糖类寡糖等含量显著增加[7]。此外越冬期间可溶性蛋白、氨基酸、活性氧清除酶等都会发生一系列适应低温的变化,Sambe等[8]研究表明抗寒性与冷诱导蛋白的积累成正相关,增强细胞抗结冰脱水能力维持细胞膜稳定进而提高抗寒性和抗冻性[9]。但目前关于越冬期间水分不同引起的干旱造成根颈脱水死亡影响返青的机制研究未见报道,由于田间不可控因素多,易受气候、风速、温度等因素的影响且不同地块苜蓿根颈失水程度不同,故本试验采取室内模拟越冬期间不同水分的低温锻炼过程,探究低温下根颈各形态、生理指标与水分之间的关系以期揭示苜蓿根颈的耐脱水机理,为科尔沁沙地地区紫花苜蓿冬春季越冬技术及当地适宜品种的筛选提供理论依据。
1 材料与方法
1.1 试验材料
2017年6月4日在中国农业大学农学院温室用花盆种植不同秋眠级的苜蓿品种(具体品种及秋眠级见表1),每品种各21盆,花盆规格为:上口径16 cm,下口径9.5 cm,高23 cm,种植基质为内蒙阿旗地区原生沙土,每盆保证出苗10株,出苗后进行正常田间管理浇水、除草等,每14 d浇灌一次营养液,温室温度控制在25~30 ℃,为保证苜蓿具有较强的生长能力,在正式低温试验之前,每35 d对苜蓿进行一次刈割以促进根系的生长发育。于2017年11月15日将试验材料刈割至8 cm并从温室移至人工气候室并于20 ℃条件下生长3 d,于2017年11月19日开始人工模拟越冬处理,在开始抗寒锻炼之前在花盆底部及外围包裹保温材料,先包裹长51 cm×宽23 cm×厚15 mm带背胶的橡塑海绵,再包裹3层长53 cm×宽30 cm×厚5 mm的毛毡布,包裹完成后开始模拟越冬试验。
1.2 试验方法
模拟越冬过程分以下阶段进行:1)抗寒锻炼期: 第1阶段将花盆置于白天10 ℃(14 h,光强为7000 Lx),夜间5 ℃(10 h)的低温气候室生长7 d,第2阶段调整温度为白天5 ℃ (14 h,光强为7000 Lx),夜间2 ℃(10 h)并生长7 d。整个抗寒锻炼过程保持土壤含水量为田间持水量的90%~95%,抗寒锻炼期结束后对每盆材料进行换土,换土深度为从表土至根颈处3 cm。换土:替换土壤的含水量为田间持水量的40%(N1)、田间持水量的65%(N2)、田间持水量的90%(N3),每个处理7次重复,换土之后转入抗寒冰冻期的培养。2)抗寒冰冻期:白天-10 ℃ (8 h,光强为2000 Lx),夜间-10 ℃ (16 h) 并生长3 d。3)冻融交替期:光照时间为8 h,光强为3000 Lx,并于10:00-13:00设置温度为4 ℃,其余时间温度为-10 ℃,生长3 d。4)缓苗期: 白天4 ℃(8 h,光强为3000 Lx),夜间4 ℃(16 h)条件下缓苗3 d。 缓苗期结束后,一部分材料用于越冬指标的测定,一部分送回温室,在温室条件下进行返青。送回温室后,各试验材料恢复正常供水即为田间持水量的75%~80%。

表1 供试品种Table 1 The experiment materials
注:数据来自北京正道生态科技有限公司、克劳沃有限公司和北京佰清源畜牧业科技发展有限公司产品宣传册。“-”表示抗寒级数未知。
Note: The data are from the brochures of Beijing Rytway seed Company Limited, Clover Company Limited and Beijing Green Animal Husbandry S&T Development Company Limited. “-”means the cold resistance is unknown.
1.3 指标测定
迅速剪取经不同水分条件越冬并缓苗之后的各试验材料根颈处 1 cm,立即用冷水冲洗并测定根颈水势,其余根颈迅速用液氮保存并存至-80 ℃冰箱,用于后续指标的测定。剩余材料去除花盆外部保温装置后送回温室进行返青观测并于14 d后测定返青率及地上部鲜重和干重。用游标卡尺测量根颈直径,根颈收缩处直径X1与最膨大处直径X2,根颈收缩比=(X2-X1)/X2×100%;采用1505D-EXP(Model 1505D Pressure Chamber Instrument,美国)气穴压力势植物水势仪测定根颈水势;采用蒽酮硫酸比色法测定可溶性糖、淀粉含量[11-12];采用酸性茚三酮比色法测定游离脯氨酸含量[13];采用戴安ICS 3000离子色谱仪(DIONEX ICS-3000,美国) 与安培检测器测定根颈蔗糖、棉子糖、水苏糖、果糖、甘露糖和半乳糖含量。
越冬后返青指标的测定,返青后统计每个处理苜蓿苗的返青率、返青枝条数、返青地上部鲜重及经105 ℃杀青15 min,65 ℃烘干至恒重的地上部干重。
1.4 数据统计分析
采用 SPSS 19.0 统计软件整理并对试验数据进行分析,采用两因素和 One-way ANOVA 进行方差分析,用Duncan法多重比较,并用 Excel 2013 制作图表。
2 结果与分析
2.1 不同水分条件越冬对各品种根颈收缩水势、可溶性糖含量及淀粉含量的影响
除WL363外,其余品种的根颈收缩比在N1、N2水分下越冬时差异不显著,但都显著高于 (P<0.05) N3条件下的根颈收缩比(图1)。N1水分下骑士T根颈收缩程度最小,为12.26%。N2水分下WL363、巨能4015根颈收缩明显。N3水分下各供试品种根颈收缩差异均不显著,WL168根颈收缩最小为5.18%。各品种在N3水分下越冬时根颈水势显著高于(P<0.05)N2和N1水分条件,N3水分条件下WL298根颈水势显著低于其他(图1)。N2、N1水分下骑士T根颈水势显著高于其他(P<0.05)。除WL363外其余供试品种根颈可溶性糖含量随水分的增加呈上升趋势,抗寒级数较大的WL168、WL298、公农一号、骑士T在不同水分下越冬后根颈可溶性糖含量之间差异不显著,而抗寒级数较小的WL363、巨能4015均表现为N1水分显著降低可溶性糖含量(P<0.05)。根颈淀粉随越冬水分增加呈下降趋势。N1水分下,抗寒级数较大的WL298淀粉显著低于 (P<0.05)其他,而抗寒级数较大的WL168淀粉含量为供试品种最大值。N2水分下抗寒级数较小的巨能4015根颈淀粉最少。N3水分下WL298淀粉含量最少(图1)。
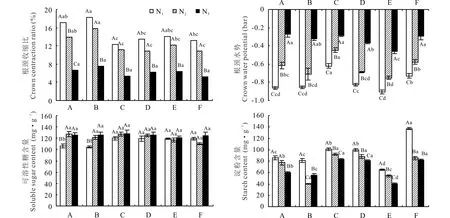
图1 各品种在不同水分条件下越冬对根颈收缩、水势、可溶性糖及淀粉的影响Fig.1 Effect of different moisture treatments on crown contraction, water potential, soluble sugar and starch during the winter of different varieties A:WL363;B:巨能4015 Magnum 4015;C:骑士T Knight T;D:公农一号Gongnong No.1;E:WL298;F:WL168.不同小写字母表示同水分不同材料差异显著(P<0.05);不同大写字母表示同材料不同水分差异显著 (P<0.05),下同。The different lowercase letters indicate significant differences in moisture at different materials (P<0.05); The different capital letters indicate that the same material is significantly different under different moisture levels (P<0.05), the same below.
2.2 不同水分越冬对各品种根颈糖分的影响
由图2可以看出,可溶性糖中蔗糖所占比例最大,最高达85%。各品种中,根颈蔗糖含量随越冬水分增加呈上升趋势,N3水分条件下WL363蔗糖含量最大,骑士T、WL298次之。N2水分条件下各品种蔗糖与N3比均显著降低。棉子糖含量随越冬期间水分含量增加呈上升趋势,N3水分条件下抗寒级数较大的WL298、WL168棉子糖含量显著高于(P<0.05)其他。对于抗寒级数较小的巨能4015,N1、N2水分条件下显著降低其根颈棉子糖含量(P<0.05)。水苏糖随越冬期间根颈水分增加呈上升趋势,抗寒级数较大的WL298在各水分条件下水苏糖含量均表现较好,N3水分条件下水苏糖含量最大。N1水分条件下WL298、WL168的水苏糖含量较高。骑士T、公农一号、WL298 和WL168 根颈中的果糖含量随越冬期间土壤水分含量的增加呈先上升后降低的趋势,抗寒级数较小的WL363、巨能4015则表现为随土壤水分含量的增加果糖含量呈下降趋势。除巨能4015、骑士T外,根颈甘露糖含量随土壤水分含量的增加呈上升趋势。N2水分条件下巨能4015、骑士T具有较高的甘露糖含量,且显著高于其他各品种(P<0.05)。根颈半乳糖含量随越冬期间土壤水分含量的增加呈上升趋势,N3水分条件下骑士T品种的根颈半乳糖含量最高。
2.3 不同水分越冬对各品种根颈脯氨酸含量及相对电导率的影响
越冬期间根颈脯氨酸含量随根颈水分增加呈上升趋势(图3),且N3水分条件下各品种脯氨酸含量显著高于N2、N1条件(P<0.05)。N3水分条件下抗寒级数较大的WL298、WL168脯氨酸含量显著高于其他(P<0.05)。随着越冬水分增加根颈相对电导率呈下降趋势,抗寒级数较大的WL168在N3水分条件下根颈相对电导率显著低于N1、N2(P<0.05),随水分增加电导率下降幅度分别为为5.9%和14.7%。对于抗寒级数较小的WL363而言,各水分处理间根颈的相对电导率差异不显著。

图2 不同水分条件下越冬对各品种根颈糖分的影响Fig.2 Effect of different moisture treatments on crown sugars during the winter of different varieties

图3 不同水分越冬对各品种根颈脯氨酸含量和相对电导率的影响Fig.3 Effect of different moisture treatments on crown proline and relative conductivity during the winter of different varieties
2.4 不同水分越冬对各品种返青性能的影响
随越冬期间土壤水分含量的增加返青率呈上升趋势,抗寒级数较大的WL168在各水分条件下返青率都表现较好,WL363、巨能4015的返青率次之,平均达73.17%。N2水分条件下WL363返青率最高达66.67%,骑士T返青最差仅35.33%。N1水分条件下WL298、WL168返青率显著高于其他(P<0.05)(图4)。此外,随着越冬期间土壤水分含量的增加,返青苜蓿的枝条数呈上升趋势,N3水分条件下各品种苜蓿的返青枝条数显著高于其他水分条件(P<0.05),骑士T的返青枝条数最高。N1水分条件下巨能4015、骑士T和公农一号的返青枝条数都显著低于WL298、WL168(P<0.05)。返青后地上部鲜重随土壤水分含量的增加呈上升趋势,N3水分条件下WL298品种的地上部鲜重显著高于其他品种(P<0.05),WL363、巨能4015的地上部鲜重最小,平均为11.71 g。N1水分条件下WL168鲜重表现最好,巨能4015在N1水分条件下鲜重最少,显著低于其他各品种(P<0.05)。返青后地上部干重随越冬期间土壤水分含量的增加呈上升趋势,N3水分条件下WL363、巨能4015的干重显著低于其他品种(P<0.05),N2水分条件下WL298、WL168的干重较大,但各品种间差异不显著。N1水分下WL298干重显著高于其他品种(P<0.05),而WL168的干重最低。

图4 不同水分越冬后各品种返青性能Fig.4 Effect of different moisture treatments on regrowth performance during the winter of different varieties
3 讨论
越冬期间干旱会造成苜蓿根颈部位失水收缩,刘志英[14]研究结果标明越冬期间秋眠级低抗寒级数大的苜蓿根颈直径较大,说明抗寒性强与苜蓿的根颈性状有一定的关系,而本试验结果表明秋眠性低抗寒级数较大的WL168、骑士T、公农一号、WL298根颈收缩都显著低于秋眠性高的WL363和巨能4015,这再次证明根颈的性状与苜蓿的抗寒性有关。王运涛[15]研究结果表明秋眠级较低苜蓿根颈粗壮表现出更强的越冬率,越冬期间根颈入土较深,根颈周围分布较多的根颈芽及分枝本研究中,抗寒性较强的WL168的分枝数在越冬后也较多,本研究结果与此一致。水势反映植物的水分状况,韩文军等[16]对阿拉善荒漠区盐生植物的日水势研究表明植物在水分胁迫下通过水势变化可反映植物体含水量的多少,从而更好地探讨植物耐逆境胁迫的机理,本试验各品种N3水分下越冬时根颈水势显著高于N1和N2条件,说明越冬期间尤其是冰冻期水分显著影响不同秋眠品种根颈的水势,水势越低越易造成根颈的失水干枯死亡,从而导致返青率降低,本试验结果标准在冰冻期,40%的田间持水量可显著降低根颈水势,结合返青率和分枝数的数据可以看出,田间含水量越低,其返青率和枝条数也越低,说明越冬期内土壤水分含量影响着苜蓿的生长情况,即在越冬季应注意苜蓿地水分的补充。而结合水势与根颈收缩比的结果可以看出,同品种不同水分处理条件下,土壤水分含量越低,根颈收缩比越高,根颈水势也越低,由此可以看出土壤水分含量主要通过影响根颈的活性,进而影响苜蓿的生长情况。
越冬期间可溶性碳水化合物作为渗透调节物质通过增加细胞液浓度降低冰点,缓冲细胞质的脱水避免蛋白质凝固变性,增强植物抗冻性。本试验通过对糖分的测定表明可溶性总糖中蔗糖含量最高且在不同水分条件下越冬下各品种差异显著,对于秋眠性低抗寒级数较大的WL298、WL168而言蔗糖含量显著增加,而WL363品种的蔗糖也出现较高含量的积累,可能是因为WL363相对来说仍属于秋眠性品种 (Fall Dormancy 4.9),高水分下越冬仍会促进根颈积累蔗糖从而提高越冬能力。韩文军等[16]、Jung等[17]、Castonguay等[18]的研究结果也表明可溶性碳水化合物的最主要部分是蔗糖,蔗糖的含量与苜蓿的抗寒性有一定的相关性。本试验中不同秋眠级苜蓿根颈可溶性糖含量随越冬水分增加而增加,对于抗寒级数较小的WL363、巨能4015根颈可溶性糖增加的更明显,对于抗寒级数较大的WL168、骑士T、公农一号、WL298品种,根颈可溶性糖含量随水分增加而增加但相互间差异不显著,说明这4个品种越冬期间糖分积累量较高而消耗的较少,表明这4个品种具有更强的抗寒性。Haagenson等[19]研究结果也表明秋眠性低抗寒级数越大的品种所积累的可溶性碳水化合物越多。Castonguay等[18]研究结果表明淀粉在越冬期间可转化成可溶性糖,从而提高苜蓿的抗寒性,本试验结果表明随着越冬期间土壤水分条件的增加,根颈中的淀粉含量呈下降趋势,而可溶性糖含量升高。Castonguay等[18]研究表明诸如棉子糖、水苏糖类的寡糖积累与苜蓿的抗寒性相关,本研究结果也表明,根颈中棉子糖、水苏糖类和寡糖的含量随越冬期间土壤水分含量的增加而增加,其中抗寒性较强的WL298、WL168、WL363苜蓿品种的增加量较大,而骑士T和巨能4015的棉子糖、水苏糖类和寡糖积累量相对较少。
脯氨酸是重要的渗透调节物质,Jaleel等[20]研究表明干旱胁迫下,植物通过调节和特化气孔结构来增强保水能力,通过促进根系生长、积累脯氨酸和可溶性糖等物质提高渗透调节能力,通过抗氧化剂和活性氧清除酶来降低活性氧伤害。本研究结果表明N3条件下WL168、WL298、WL363积累的脯氨酸最多,而干旱条件显著降低各秋眠品种的脯氨酸含量,这与Dhont等[21]的研究结果相似,说明脯氨酸可能对越冬时水分影响较敏感,水分降低会导致各品种脯氨酸的降低影响根颈抗寒性和苜蓿抗冻性。Sule等[22]研究表明,抗寒性强的苜蓿品种细胞内溶物泄漏率低,冷冻对其造成的伤害较小。本研究结果表明,相对电导率随越冬水分增加呈下降趋势,说明抗寒级数较大的品种受离子胁迫程度小,抗寒级数较小的WL363、巨能4015各处理间相对电导率差异不显著,抗寒级数较小的品种受水分影响较大,这与它们本身的秋眠性相对较高有关。
越冬后的返青率及返青枝条再生数、返青后株高、鲜重、干重等都能代表一个品种越冬率及抗寒性的强弱。因此对越冬后的返青数据采集能更全面地反映一个品种的抗寒性,本研究结果显示返青率与秋眠性相关,秋眠性越低抗寒级数越大的品种返青率越高。返青枝条数随越冬水分增加显著增加,说明越冬期间水分对返青率产生影响,相比于抗寒级数较大的WL298、WL168品种,巨能4015返青率及返青枝条数、鲜重都显著受到抑制,说明品种的抗寒级数还是影响越冬及产量的因素之一,WL363虽秋眠性也高但可能由于越冬期间积累较多的脯氨酸、蔗糖、棉子糖等表现出较高的返青率及枝条数,也可能是由于打破越冬休眠后刺激WL363根颈促进再生枝条的萌发和生长,也有可能是 WL363 再生节点低、再生性强越冬恢复性较强,有待进一步开展试验进行探究验证。孙予璐等[23]研究结果表明抗寒性强的品种低温胁迫下积累较高的可溶性糖从而提高苜蓿的抗寒性,而本试验也证实抗寒级数较大的 WL298、WL168 经不同水分越冬并返青后返青率及返青枝条数、返青后干鲜重均表现最佳,说明 WL298、WL168 根颈耐脱水较强,即在较低的水分条件下根颈也能完成越冬保护性糖的生理进程,减少越冬伤害。
4 结论
1)模拟越冬季不同水分显著影响各品种根颈越冬保护性糖的积累及根颈的生理活性,尤其是蔗糖、棉子糖、水苏糖类寡糖及果糖的积累。
2)抗寒级数较大的WL298、WL168根颈耐脱水较强。骑士T、公农一号根颈耐脱水性中等。而抗寒级数较小的巨能4015根颈耐脱水性较弱。WL363虽秋眠级高但由于积累越冬保护性糖的含量高、根颈耐脱水能力及再生恢复性较强表现出较强的越冬能力,根颈耐脱水性中等。
3)越冬水分维持在田间持水量90%时,WL298、WL168、骑士T、公农一号的越冬保护性糖及返青后生产性能均强于巨能4015。越冬水分维持在田间持水量40%及以下各品种根颈收缩程度显著增加,WL298、WL168、WL363返青性能较好而骑士T、公农一号和巨能4015则返青率相对较低。
