人为采笋对大叶筇竹生长及大熊猫采食的影响
2019-09-21白文科王光磊杨旭煜古晓东周材权
白文科, 董 鑫,王光磊,杨旭煜,古晓东,周材权
(1.西南野生动植物资源保护教育部重点实验室,四川 南充 637002;2.西华师范大学生态研究院,四川 南充 637002;3.四川省野生动物资源调查保护管理站,四川 成都 610081)
大熊猫(Aihuropiodamelanoleuca)是我国特有的珍稀濒危物种,具有十分重要的科学研究价值,在学术界更被誉为“活化石”[1~2]。大熊猫为食肉目动物,随着环境的改变,经过数千年的演化,逐渐演化成以竹类为主食的植食性动物[3]。作为专食性动物,大熊猫虽然保留着食肉动物的消化道,但竹子在大熊猫的食谱中却占到了99%以上[4]。
在马边大风顶自然保护区,20世纪90年代曾对大熊猫种群数量[5]、栖息地选择[6]、主食竹等方面做了大量的研究[7-9]。近20年来,马边大熊猫数量急剧下降,1977年全国大熊猫第一次调查时,在保护区范围内发现大熊猫36只。1988年全国第二次大熊猫调查时,保护区的大熊猫数量为32只。1991—1992年间,马边大熊猫数量为36 ± 6只[5]。到2001年全国第三次大熊猫普查时,马边大熊猫的数量下降到11只。短短不到10年时间,马边大熊猫数量减少20多只,而在之后的10年大熊猫的数量有所回升,2012年全国第四次大熊猫调查结果显示大熊猫数量达到了18只[10]。
大叶筇竹(Qiongzhueamacrophylla)为无性系种群,采笋之后无性系种群能够进行自我调控,使地下茎更多的萌发出新笋来弥补子代分株数量的不足。大叶筇竹具有重要的生态、经济价值,竹秆秆环隆起较大,经常被做成手杖和工艺品,竹笋味甘鲜嫩,具有较高的经济价值[11],是人为采笋的主要竹种之一。同时,人为采笋是大熊猫栖息地主要的干扰因子,过度采笋活动不仅使得采笋后剩余竹笋量明显下降,还会促使新笋不断萌发,这是竹林采取投资克隆繁殖的能量相对较多的生态策略[12],但由于营养缺乏,导致产生的新竹笋数量多而质量较差,进而造成大叶筇竹生长状况明显下降[13]。
在大熊猫栖息地内,主食竹的生长状况是大熊猫存活和生境选择的重要影响因素之一,1974年和1983年,大熊猫栖息地出现箭竹大面积开花死亡现象,就造成了约250只野生大熊猫由于主食竹短缺而死亡的严重后果[14]。因此,对大熊猫栖息地人为采笋活动和主食竹的生长状况等方面进行调查,进而分析栖息地变化情况,为更加科学、合理的保护自然资源,更好的保护大熊猫这一濒危物种提供理论依据。
1 材料与方法
1.1 研究区概况
四川马边大风顶自然保护区位于四川省西南部马边彝族自治县境西南边缘,位于东经103°13′~103°26′和北纬28°26′~28°45′之间。地处四川盆地和云贵高原的过渡地带,青藏高原的东南缘,横断山脉中段,属全球生物多样性地区之一的中国西南山地地区。保护区建于1978年,是以保护大熊猫(Ailuopodamelanolcuca)、羚牛(Budorcastaxicolor)、珙桐(Davidiainvolucrata)、水青树(Tetracentronsinense)等珍稀濒危野生动植物及其生境为主的野生动物和森林类型自然保护区。保护区内植被分带现象较为明显,植被垂直带谱结构较完整,海拔 1 300 m~1 800 m分布有低山常绿阔叶林,林下竹类为小叶筇竹(Qiongzhuegtumidinoda)、刺竹(Chinmonobambusapachstachys)、四川方竹(Chimoncdambusaszechuanensis);海拔1 800 m~2 200 m之间为常绿与落叶阔叶混交林,林下竹类为大叶筇竹(Qiongzhueamacropohylla);海拔2 200 m~2 500 m为针阔混交林,林下竹类为大叶筇竹和刺竹;海拔 2 500 m~3 500 m为亚高山针叶林,林下竹类为马边玉山竹(Yushaniamabianensis)和冷箭竹(Bashaniafangiana)。马边大风顶自然保护区竹林分布广泛并且种类较多,春、秋两季都有竹林发笋,主要集中在每年的4月和9月,为大熊猫提供了丰富的食物资源。
1.2 研究方法
1.2.1 野外数据搜集
马边大风顶自然保护区周边居民一直延续采笋的传统,随着采笋收入及占比的逐年增加,保护区及周边区域的人为采笋强度也增加明显。本次研究以户为单位,在采笋季节对笋农进行了走访调查,对笋农组成及收入进行了统计。并于2013年5月在采笋区域内避开历史森林砍伐区,选择与1993年数据搜集区域相似的坡体,设定随机样线收集大熊猫在大叶筇竹林内的活动痕迹,并沿随机样线设置竹子样方,样方大小为1 m × 1 m,间隔距离为50 m,统计大叶筇竹数量,并使用卷尺和游标卡尺分别测量样方内所有大叶筇竹立竹的基径和高度。
在调查区大叶筇竹林分布区内,结合地形图、植被图以及保护区大熊猫监测样线调查数据,同时在笋农走访调查中咨询笋农,划定大熊猫在大叶筇竹林区域内的重点调查区。最终,共设置固定样线6条,收集大熊猫在大叶筇竹林内的活动痕迹。并与多方面调查(保护区大熊猫监测样线、大熊猫种群数量调查、大熊猫交配场所调查)结合,分析大熊猫对大叶筇竹的利用现状。
1.2.2 室内数据分析
在时间跨度上,将2013年调查数据与1993年相同区域大叶筇竹的生长状况数据进行比较分析,1993年数据来源于本文作者在马边大风顶自然保护区大熊猫观察站调查的数据统计。使用Spss 22.0中的独立样本T检验(Independent-Sample T Test)进行统计数据的差异分析。
2 结果与分析
2.1 走访调查结果
本次共调查笋农49户,合计参与采笋249人。采笋人员中女性占62.6%,远高于男性,主要是很多男性外出打工,而女性受教育程度低,很多不会汉语,外出很不方便,加上笋期农活较少,因此多数妇女可在笋期进山采笋。在人员年龄组成上,主要以青壮年为主,但也有未成年和老年人也进山采笋,在调查中最高的笋农年龄为71岁。采笋人员受教育程度普遍较低,初中及以上教育程度的只有11人(其中两人为高中教育),仅占调查人数的4.5%。以户为单位取平均值估算了笋农家庭的收入组成,年均家庭收入为 11 090.62元,其中采笋收入为 4 730.15 元·a-1,占家庭年总收入的比重较大,达到42.65%;人均采笋收入为927.48元·a-1,是保护区周边居民的主要经济来源之一。
2.2 人为采笋对大叶筇竹生长状况的影响
2.2.1 对大叶筇竹密度的影响
与1993年大叶筇竹长势相比,2013年调查时大叶筇竹的密度下降明显(P<0.05),竹子密度由17.30株·m-2下降到了12.94株·m-2;采笋强度增加后竹笋量下降明显(P<0.05),竹笋量由4.34株·m-2下降到2.32株·m-2;而竹子死亡数也有略微下降,但差异性不显著(见表1)。大叶筇竹林竹子密度的差异表明:人为采笋活动使得大叶筇竹林的竹子密度和竹笋量受到了显著的影响,两者均出现了明显的降低。
表1大叶筇竹密度的变化

Tab.1 The change of density of bamboo
2.2.2 对大叶筇竹基径和高度的影响
与1993年大叶筇竹长势相比,2013年调查时大叶筇竹的平均基径下降明显(P<0.05),基径均值由11.61 mm下降到10.63 mm;大叶筇竹的平均高度略有下降,但差异性不显著(P>0.05),高度均值变化不大(见表2)。竹林竹子基径的差异表明:人为采笋活动使得大叶筇竹林的基径受到了显著的影响,出现了明显的降低;但对竹子高度的影响不明显。
表2大叶筇竹基径和高度的变化
Tab.2 Thechangeofbasaldiameterandheightofbamboo
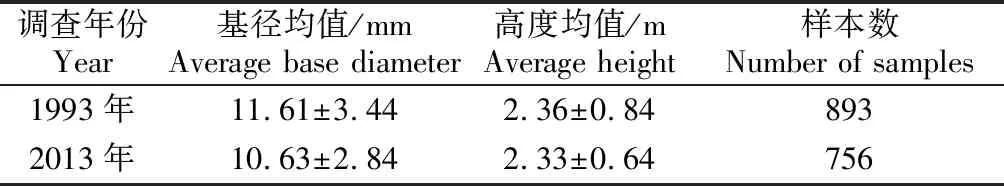
调查年份Year基径均值/mmAverage base diameter高度均值/mAverage height样本数Number of samples1993年11.61±3.442.36±0.848932013年10.63±2.842.33±0.64756
2.3 人为采笋对大熊猫采食的影响
统计了在大叶筇竹林分布区2013年3—11月的调查结果,只在4月份发现两团全部以大叶筇竹竹叶为食的大熊猫粪便,该粪便新鲜程度时间在一个月左右,表示大熊猫已很少在大叶筇竹林中栖息采食。在大熊猫交配场的调查中,共发现两处交配场,收集到大熊猫新鲜粪便24团,全部位于海拔更高的马边玉山竹(YushaniadafengclingensisYi)分布区。调查结果也可以证实,马边大风顶自然保护区的大熊猫现阶段主要以不采笋的马边玉山竹为食,几乎不以大叶筇竹为食。同时走访调查还发现,刺竹子分布的海拔更低,人为活动更为严重,大熊猫几年内都不利用刺竹林分布区域。与之前的调查结果相比,人为采笋活动已经影响到了大熊猫食性的选择。
3 讨论
马边大风顶保护区周边居民一直延续着采笋的传统,20世纪70年代以前的采笋主要是以自己食用为目的,这一时期的采笋范围较小,主要集中在低海拔的区域。但随着竹笋价格的上涨,参与竹笋采集的人群、采笋的数量以及采笋的范围都大幅度增加。在1993—1997年的森林砍伐期间,保护区内修设的运木道路为保护区周边笋农的采笋活动提供了便利,使笋农的采笋活动范围进一步扩大。另外,据当地笋农介绍:保护区周边属山地地貌,耕地多分布在山坡上,每年春季居民种完庄稼后,在春笋时期(4月—6月),因家中农活较少,几乎家中所有的劳动力都进山采笋;而秋笋时期(9月—10月),因家中农活较多,需要留一部分劳动力在家务农,因此秋季采笋人员要相对少一些。但是,由于每个采笋期过后,笋农都需要把晾干的竹笋不定期地运送到山下,所以在每年的4月—11月一直有笋农频繁地活动在大熊猫的理想栖息地内。
1993年政府在保护区内砍伐森林、修设运木公路[15],使马边地区大熊猫栖息地受到的人为干扰达到了最大化。但不同强度的采伐对大熊猫栖息地的植物群落组成、结构和林下竹类的生长质量有较大的影响,森林砍伐一定程度上增加了土壤中微生物种群数量,适度的森林砍伐更有利于栖息地竹类质量的提高[16]。马边自然保护区内的森林砍伐在一定程度上也改变了大叶筇竹林内的竹子结构和竹子的生长状况,森林砍伐导致上层乔木减少,森林郁闭度下降,适合大熊猫栖息地选择的生境大幅减少,人为砍伐过后,生境的物种优势度、群落高度和最大树径明显下降,这对大熊猫栖息地的选择产生影响[17]。大熊猫在栖息地的选择上,更喜欢选择上层乔木郁闭度较大(大于50%)的大叶筇竹林中觅食[7]。因此,虽然有生长状况良好的大叶筇竹,但也使得大熊猫逃离了这一区域,在栖息地空间上受到了严重挤压,而且森林砍伐时修的运木道路,某种程度上也扩大了采笋活动的范围和采笋强度,使人为采笋活动对大叶筇竹的干扰进一步增大。
魏辅文等对马边大熊猫食性的研究发现,马边大熊猫喜食基径较粗(>10 mm)、幼竹比例较高(>5%)、密度适中(10株·m-2~30株·m-2)的大叶筇竹[6]。长期的采笋活动导致大叶筇竹基径、幼竹比例、高度和竹子密度均下降明显,采笋过后的剩余竹笋下降显著,竹笋比例严重下降,竹子死亡数略微下降,进而导致马边大熊猫几乎难以大叶筇竹为食,而被迫以以前很少采食的白背玉山竹(YushaniaglaucaYi et T.L.long)为食,食物资源质量下降也有可能导致大熊猫对大叶筇竹林的利用率下降,以至于人类的采笋活动几乎已经影响到了大熊猫食性的选择。20世纪90年代初就有研究显示:在未受干扰之前,马边大熊猫冬季的活动痕迹大部分都在海拔 2 100 m~2 400 m 的大叶筇竹林里,而5月发笋后,采笋活动的干扰影响会使大熊猫的活动范围上移至海拔 2 400 m之上,而在禁止采笋后则会迅速下移,大熊猫会因为采笋活动的影响而推迟采食竹笋的时间[10]。
采笋活动不仅导致大叶筇竹质量和幼竹比例下降,更主要的影响因素是人为活动时间长,从4月—6月、9月—10月,并且笋期结束后,鲜笋要在山上放干后再运下山,这就导致几乎从4月到11月保护区内一直有人活动。因此,采笋活动对大熊猫生境选择的影响不仅体现在对大熊猫食物的掠夺上,也体现在人类的大范围持续采笋活动干扰上,各种干扰迫使大熊猫离开了人类采笋活动的区域。另外,人类的过度采笋不仅破坏大熊猫食性选择规律和活动规律,更重要的是大量的人员涌入山林,难免不对大熊猫造成伤害,据报道,仅1993年4月—7月份的采笋季节,马边县境内共发现大熊猫尸体3只,造成其中至少两只大熊猫中毒死亡的误毒事件[14]。
在马边县大风顶自然保护区及其周边区域的人为采笋已经影响到马边大熊猫赖以生存的栖息地和食物资源,而周边社区对保护区资源的索取也是生活必不可少的一部分。马边大风顶自然保护区管理局已经在保护区内实行了轮息采笋等相关保护政策,同时随着社会经济的发展,保护区周边居民收入来源有所增加,采笋的程度已有所下降。但保护区范围外的采笋强度依然很大,对竹林资源的健康发展产生了巨大的压力。为了更有效的保护大熊猫以及更合理地利用保护区资源,势必采取有效措施,改善周边社区生活条件,减少对保护区及其周边自然资源的压力。
