土壤动物群落对桢楠人工林边缘效应的响应
2019-09-21罗熳丽涂程伟辜云杰
王 戈,罗熳丽,张 亚,涂程伟,辜云杰
(1.四川省九顶山省级自然保护区,四川 什邡 618400;2.四川农业大学林学院,四川 成都 611130;3.四川省林业科学研究院,四川 成都 610081)
土壤动物是土壤生态系统的重要组成部分,作为分解者参与土壤生态系统的物质和能量循环,是森林土壤肥力的重要指标[1~4]。土壤动物群落和植物的互动影响着生态系统的过程[5~6],而土壤生态系统又是森林生态系统不可或缺的一部分。因此,土壤动物对生态系统的功能和稳定性具有重要意义[7~11],并能在一定程度上指示环境的变化[12~15],如生境的变化,因此对于不同生态系统下的土壤动物群落结构[16]、生态功能[7]及多样性[11]研究普遍,但对于边缘效应对土壤动物的影响研究较少。
边缘效应是指在两个或多个不同性质的生态系统(或其他系统)交互作用处,由于某些生态因子(可能是物质、能量、信息或地域)或系统属性的差异和协和作用而引起系统某些组分及行为(如种群密度、生产力、物种多样性等)的较大变化[17]。边缘效应是生态学和生物保护的重要概念之一,它在研究生态系统尺度和景观生态系统尺度的能量流和物质流等生态过程中具有重要的作用[18]。目前针对边缘效应对植被的研究内容已经比较丰富,但是针对边缘效应对土壤动物群落特征的研究报道还比较少。本研究以人工桢楠林为例,探讨边缘效应对土壤动物群落结构特征影响以及研究方向,以期为森林经营、保护区管理等生产实践和生态环境综合治理提供科学依据。
1 研究样地与研究方法
1.1 试验地概况
样地设在成都平原与四川盆周西缘山地接合部的都江堰灵岩山(103°25′42″~103°47′E,30°44′54″~31°22′9″N)58年生的桢楠人工林纯林,样地土壤为砂岩上发育的黄壤,质地为重壤质[18]。pH值6.5~6.8。由于多雨,在淀积层与母质层之间有明显的潜育现象,土壤肥力中等,保肥保水性好。海拔780 m~1 009 m,年平均气温16 ℃,年均最低气温5 ℃,年均最高气温35 ℃[19],平均年降水量为 1 225.4 mm,年平均日照时数 1 024.2 h。
1.2 研究方法
1.2.1 实验地设置
于2016年8月,在四川省都江堰市灵岩山人工桢楠林选择较少受到人为干扰的地块作为调查区域。按照不同的种植密度,采用随机抽样的方法,共设置两个大小为10 m×10 m的样地,2个宽度为10 m的样带。
1.2.2 实验方法
于2016年8月分别对所设样地和样带进行土壤动物群落调查和分析。在林地中心的各样地内分别按“品”字型布点,在林地边缘的各样带设置3个50 cm×50 cm(0.25 m2)的样方,土壤剖面分凋落物层、0 cm~5 cm、5 cm~10 cm、10 cm~15 cm 共4个层次,并对其进行手捡,将所得的土壤动物放入75%浓度的酒精容器中杀死,按样地、样方注明编号,带回实验室在解剖镜下观察鉴定;在每个样地内采3份10 cm×10 cm(0.01 m2)枯落物,用尼龙网包好,编号并放入黑布袋,带回实验室对土壤动物进行分离;另外,在各样点挖土壤剖面,分0 cm~5 cm、5 cm~10 cm、10 cm~15 cm 共3个土层,各土层用圆形取样环刀(r=5 cm,v=100 cm3)自上而下取土,每层各取两个,用尼龙网包好土样,编号后放入黑布袋,完整带回实验室用Tullgren干漏斗(干生)和Baermann湿漏斗(湿生)对土样中的土壤动物进行分离。土壤动物的分离参照黄玉梅[1, 5, 21]等的方法,土壤动物的分离均在烘虫箱中进行,烘虫箱的温度控制在35℃~40℃。干生、湿生、枯落物的分离时间均是48 h,其中湿生4 h观察1次,分离出的土壤动物除湿生以外,都用盛有75 %浓度的酒精培养皿收集,在解剖镜下观察鉴定土壤动物种类并计数,湿生土壤动物的收集用清水,最后在解剖镜下进行分类鉴定和数量统计。
1.3 土壤动物鉴定
土壤动物的分类鉴定:将手捡和分离所得的土壤动物放置在双目解剖镜(Leica,EZ4HD)下观察;主要采用《中国土壤动物检索图鉴》[22]、《中国亚热带土壤动物》[23]和《昆虫分类检索》[24]进行分类鉴定,一般鉴定到科的水平。
1.4 多样性指数
类群等级划分:个体数量占捕获总量的10.0 %以上者为优势类群(+++), 在1.0 %~10.0 %为常见类群(++), 小于1.0 %者为稀有类群(+)[20]。
群落的多样性分析:土壤动物多样性主要采用Shannon-Wiener多样性指数H′、Margalef丰富度指数D、Pieluo均匀度指数J和Simpson优势度指数C来计算[24, 25]。
Shannon-Wiener多样性指数(H′):
(1)
式中:Pi=ni/N,ni为第i个类群的个体数;N为所有类群的个体数。
Margalef丰富度指数(D):
D=(S-1)/lnN
(2)
式中:S为类群数,N为所有类群的个体数。
Pieluo均匀度指数(J):
J=H′/lns
(3)
式中:H′为Shannon-Wiener多样性指数,s为类群数。
Simpson优势度指数(C):
(4)
式中:ni为第i个类群的个体数;N为所有类群的个体数。
1.5 数据处理
差异显著性分析:采用独立样本T检验(Independent-Samples T Test)对不同样地间土壤动物群落组成进行检验,显著性水平设定为P= 0.05。
数据的整理与分析采用Excel 2010和SPSS 20.0,采用Excel 2010绘制图形。
2 结果与分析
2.1 土壤动物的组成
本次调查共捕获土壤动物887只,隶属于3门9纲12目23科(见表1)。优势类群为线虫纲,占总密度的80.30%;常见类群为丽甲螨群、绥螨科、蚁科、双翅目幼虫,分别占总密度的7.05%、4.30%、2.94%、1.20%;其他种类构成稀有类群,其密度占总密度的4.21%。在林中样地共捕获3门8纲11目17科310只土壤动物,平均密度2.69×104只·m-2;林地边缘样地中捕获3门8纲12目21科577只土壤动物,平均密度5.37×104只·m-2;林地边缘的土壤动物多,林地中心的土壤动物较少。
表1土壤动物群落组成统计
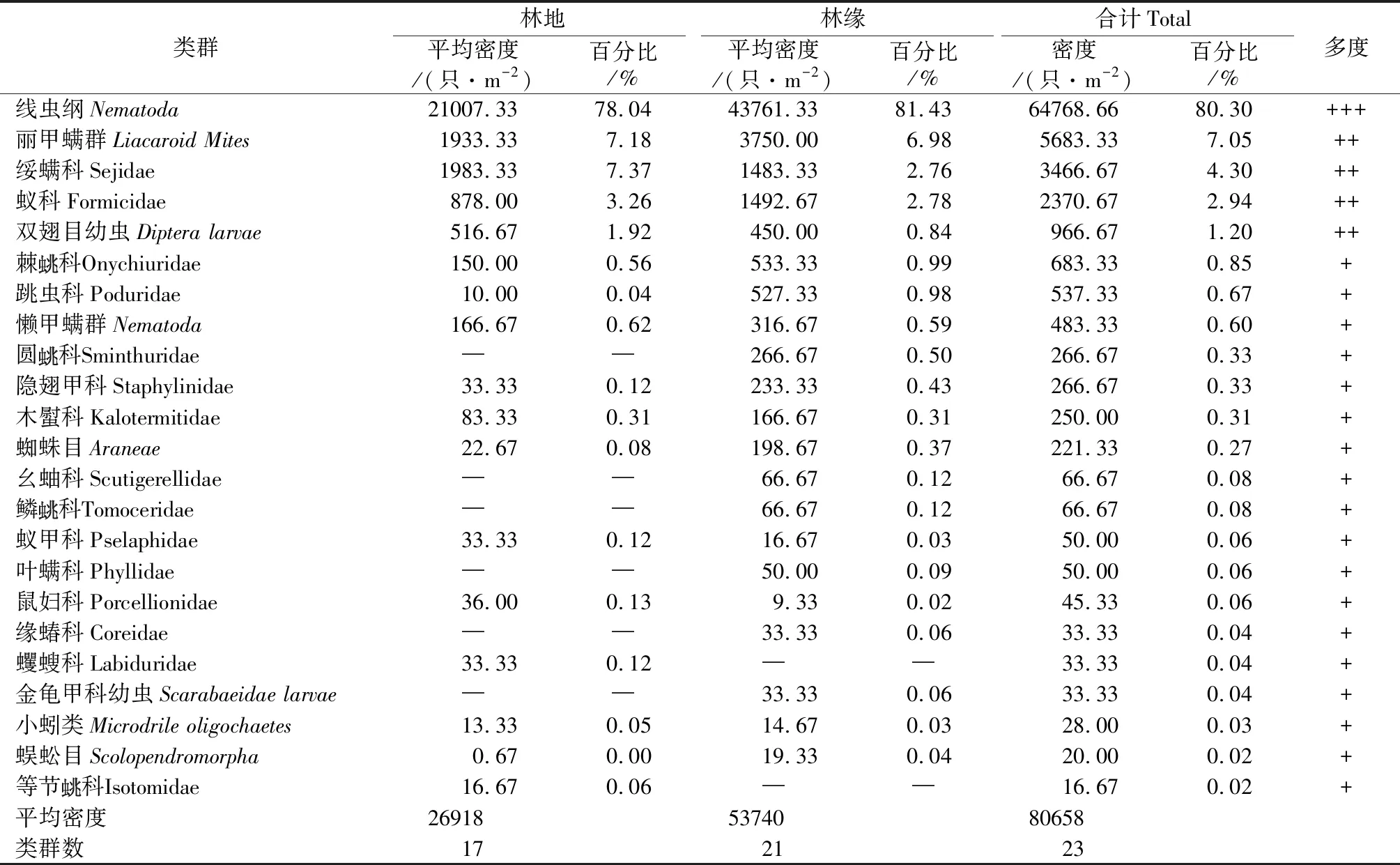
Tab.1 Compositions of soil fauna communities in the three plots
注:“─”表示无或未采集到,+++为优势类群,++为常见类群,+为稀有类群。
2.2 土壤动物群落的垂直分布特征
林地中心土壤剖面凋落物层、0 cm~5 cm、5 cm~10 cm、10 cm~15 cm层土壤动物占总土壤动物平均密度3.90%、70.82%、15.01%和10.27%;林地边缘凋落物层、0 cm~5 cm、5 cm~10 cm、10 cm~15 cm层土壤动物占总土壤动物平均密度5.14%、48.65%、21.51%和24.70%;可以看出各样地的枯落物层占比重最少,而土壤表层土壤动物比重最高(见图1)。
林地边缘样带中土壤动物的平均密度各层均高于林地中心,其中,两种类型样地土壤动物的平均密度均表现为表层土壤具有较高的平均密度,且凋落物层的土壤动物平均密度低于土壤层,土壤动物集中分布于0~5 cm层。林中样地的土壤动物群落的分布特征为:从地表向下,随土层深度的增加,土壤动物平均密度逐渐下降;林地边缘的样地的土壤动物群落的分布特征为:从地表向下,随土层深度的增加,土壤动物平均密度先降低后增大。
差异性检验结果显示:凋落物层各样地平均密度差异达到显著水平(F=3.147,P=0.016),类群数差异达到极显著水平(F=6.400,P=0.006);0~5 cm各样地平均密度差异不显著(P>0.05),类群数差异不显著(P>0.05);5~10 cm层各样地平均密度差异达到极显著水平(F=1.000,P=0.001),类群数差异不显著(P>0.05);10~ 15cm层各样地类群数差异达到显著水平(F=12.157,P=0.034),类群数差异达到显著水平(F=1.231,P=0.023)。
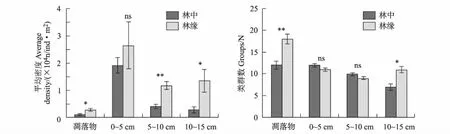
图1 各生境土壤动物垂直分布变化Fig.1 Each habitat change the vertical distribution of soil fauna注:ns表示不存在显著相关性(P > 0.05);*表示存在显著相关性(P < 0.05);**表示存在极显著相关性(P < 0.01)
2.3 干生、湿生土壤动物的变化
在两种类型的样地土壤,通过不同分离方法获得不同样地间土壤动物平均密度的存在差异,主要是干生和湿生两种分离(0 cm~5 cm层、5 cm~10 cm层与10 cm~15 cm层的土壤动物)(见表2)。林地中心土壤通过干、湿生分离出来的土壤动物分别占林地中心土壤动物密度(只含从土壤中分离出来的数量,不含手捡和枯落物中的土壤动物,下同。)5.56%、94. 4%;林地边缘土壤通过干、湿生分离出来的土壤动物分别占林地边缘土壤动物密度10.68 %、89.32 %。可以看出各样地通过湿生分离的土壤动物比干生多。
差异性检验结果显示:干生土壤动物各样地间差异达到显著水平(F=0.355,P=0.018);湿生土壤动物各样地间差异不显著(P>0.05)。在采样地点和土壤的热容量相同的条件下,土壤动物的分离效果除了受不同分离方法的影响以外,还主要受分离器内的温度影响[27, 28]。
表2各生境干、湿分离土壤动物的群落结构特征(平均值±标准误)
Tab.2 CommunitystructurecharacteristicsofeachhabitatTullgrenandBaermannseparationofsoilfauna

样地类型林地林缘干生2 866.67±1 673.65a11 000.00±1 258.31b湿生48 666.67±29 717.19a92 000.00±29 143.32a
注:同行相同小写字母表示差异不显著(P>0.05);同行不同小写字母表示差异显著(P<0.05)。
2.4 不同体型土壤动物群落的水平分布特征
按照尹文英[22]对土壤动物体型的划分,将本次调查到的土壤动物分为大型和中小型两大类。林地中心大型、中小型土壤动物分别占林地中心样地土壤动物密度的3.90%、96.10%,林地边缘大型、中小型土壤动物分别占林地边缘样带土壤动物密度的5.14%、94.86%。可以看出各样地的中小型土壤动物比大型土壤动物数量多。
林地边缘的大型和中小型土壤动物平均密度均高于林地中心(见表3)。差异性检验结果显示:两种类型样地大型土壤动物差异显著(F=3.147,P=0.016),中小型土壤动物差异显著(F=9.976,P=0.026)。
表3各生境不同体型土壤动物水平分布变化(平均值±标准误)
Tab.3 CommunitystructurecharacteristicsofeachhabitatTullgrenandBaermannseparationofsoilfauna(mean±SE)

样地类型林地林缘大型土壤动物1050.00±301.938a2800.00±527.889b中小型土壤动物25868.00±3219.098a51640.00±9365.823b
注:同行不同小写字母表示差异显著(P<0.05),不同大写字母表示差异极显著(P<0.01)。
2.5 土壤动物的多样性变化特征
土壤动物多样性是20世纪90年代以来土壤与生态系统研究的热点领域范畴。林地边缘土壤动物除均匀度指数外,其他指数均高于林地中心土壤动物。
差异性检验结果显示:两种类型的样地中Shannon-Wiener多样性指数、Pielou均匀度指数、Simpson优势度指数表现为各样地间均无显著性差异(P>0.05);Margalef丰富度指数表现为各样地存在极显著差异(F=0.231,P=0.010)。
表4各生境土壤动物群落的多样性特征(平均值±标准误)
Tab.4 Characteristicsofeachhabitatdiversityofsoilfauna(mean±SE)
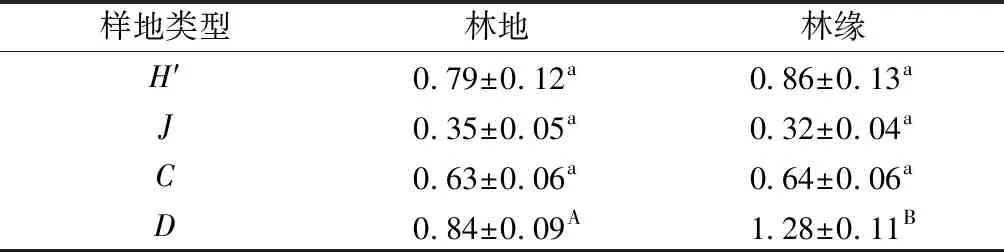
样地类型林地林缘 H′0.79±0.12a0.86±0.13aJ0.35±0.05a0.32±0.04aC0.63±0.06a0.64±0.06aD0.84±0.09A1.28±0.11B
注:同行不同小写字母表示差异显著(P< 0.05),不同大写字母表示差异极显著(P< 0.01)。
3 讨论
本次调查结果显示:桢楠人工林土壤动物群落表现出了明显的正效应,即效应区比相邻的群落具有更为优良的特性,例如物种多样性增加[29]。林地边缘的土壤动物数量和类群数均高于林地中心,产生这种影响的主要原因有:第一,内在因素的影响。对于林地中心样地来说,桢楠是常绿大乔木,叶型较小,凋落物极少,加之林地中心土壤板结现象严重,没能给土壤动物提供适宜的生存环境,导致桢楠林地中心的土壤动物在多方面指数远不及林地边缘的土壤动物。第二,外在因素的影响。核心区的林地,由于有边缘地区树木的保护,可以避免外来因子的影响,但在边缘地区,由于缺乏高大林木的保护,光照、风沙等可以直接侵入林下,从而导致林地边缘地区的生境与林地内部的生境(气温、地温、空气相对湿度、光照度等)有较大差异[18]。第三,人为因素的影响。边缘的林地因人为需要会将凋落物清扫堆积在边缘林地中,凋落物层较厚,土壤层也更为肥沃,而凋落物在森林生态系统中占有极其重要的地位,它的分解是森林生态系统养分循环的一个重要环节,不但是土壤动物最直接的物质和能量来源,其数量和种类对土壤动物的群落组成、多样性和丰富度等都有极大程度的影响[17]。
边缘效应对中小型湿生土壤动物——线虫纲造成的影响较其他土壤动物小,占总密度的80.30%,优势度较其他类群始终较高;林地边缘的土壤动物数量和类群多,林地中心的土壤动物数量和类群较少。其原因可能是由于林地边缘的凋落物层较厚,土壤腐殖质、有机质含量高,土壤孔隙度较大,利于土壤动物生存和生活。土壤动物水平变化中,各样地间大、中小型土壤动物平均密度均具有显著差异(P<0.05),其原因可能是大型土壤动物可以在凋落物和土壤不同层次间穿梭密度不稳定,而中小型土壤动物大多为腐食性,其食源主要为凋落物[1],平均密度随凋落物的变化而变化。土壤动物垂直变化中,林地边缘样地中各层土壤动物的平均密度均高于林地中心,两种类型样地土壤动物的平均密度均表现为表层土壤具有较高的平均密度,且凋落物层的土壤动物平均密度低于土壤层,土壤动物集中分布于0~5 cm层。其原因可能是由于林地中心的凋落物少而干燥、土壤板结现象严重,而0~5 cm层位于凋落物下,受到保护,土壤相对湿润,土壤质地也较为松软,适合土壤动物存活。
土壤动物的变化与森林生态系统的能量物质循环、平衡和稳定性具有很大程度的关联[30]。在桢楠人工林下,林分单一,植物功能群多样性较低,研究表明,多样性较低的林分,其土壤动物多样性、森林土壤肥力远低于多样性高的林分,以致于其生态系统功能难以维系,不利于森林可持续发展。边缘效应是自然界和人类生态系统的普遍规律,因此,必须充分利用边缘规律为整个生物圈服务。首先是开拓边缘,重视整体格局,从生物多样性和生态系统功能的角度出发,促进人工林结构优化、合理经营规划和可持续发展,提高植物的多样性,增加凋落物种类、层次和数量。通过地上和地下生物的互动,提高土壤动物多样性和丰富度,促进土壤养分元素的循环,从而使人工楠木林整个生态系统的各项功能实现可持续发展的目标。其次要调控边缘,根据实际情况的需要来对边缘予以调控。边缘地带常常是不稳定的,要维持或避免高的边缘效应,重要的是要保持或消除边缘的动态特征,从而更有利于边缘效应的利用、管理和生物多样性的保护。本实验主要初步调查了楠木人工林林中和林缘的土壤动物与土壤动物群落结构特征,揭示林地中心与林地边缘的土壤动物差异,而边缘效应的利用、管理和生物多样性的保护还有待进一步研究。
