龙岩市239株沙门菌毒力基因检测结果分析
2019-09-19曹春远陈前进陈小东钟叶平李美华陈海滨
曹春远,陈前进,陈小东,钟叶平,李美华,陈海滨
沙门菌是最重要的人兽共患传染性疾病的病原菌之一,大多数有很强的致病力,对人类健康和禽畜养殖业造成重大危害,并且是最重要的人类食源性致病菌之一,是目前的重大公共卫生问题之一。研究表明沙门菌致病主要是因为大量毒力因子相互作用导致[1],毒力相关的基因因此成为近年研究的重点。毒力基因作为沙门菌致病特性的遗传物质,也长期在发生着变迁进化。为了解龙岩市沙门菌毒力基因携带与变迁情况,本文收集了1991-2017年间239株沙门菌进行9种毒力基因检测分析,为龙岩市沙门菌病的防制提供理论依据。
1 材料与方法
1.1 材 料
1.1.1菌株来源 收集1991-2017年239株沙门菌。其中,179株分离自人体样本(主要从本市综合医院临床病人粪便样本分离,少部分分离自饮食从业人员肛拭样本),60株分离自禽畜肉等食品监测样本。
1.1.2主要仪器与试剂 VITEK 2 Compact(法国梅里埃),PCR扩增仪(德国Biometra),三恒多用电泳仪(北京六一仪器厂),凝胶成像仪(法国)。泰国S&A公司沙门菌分型血清,premix、100 bp MARKER为TAKARA产品。
1.2 方 法
1.2.1菌株复核鉴定 用VITEK 2 Compact GN鉴定卡进行复核鉴定。血清分型参照Kauffman- White标准,其中鞭毛诱导实验参照GB 4789.5-2016方法。
1.2.2毒力基因检测 用PCR方法检测沙门菌9个毒力基因的片段,包括:sopB、invA、sifA、sscA、sseE、spvB、pefA、spvR、spvC等,引物序列与扩增程序和体系配制参考文献[2-3],见表1。引物序列由上海生工合成。核酸提取应用水煮沸法。
表1 9个毒力基因片段检测用引物序列
Tab.1 Primer sequences for detection of 9 virulence gene fragments

引物名称位置引物序列5′-3′扩增片段长度来自文献invASPI-1F:TATCGCCACGTTCGGGCAATCTTR:TCGCACCGTCAAAGGAACCACT275[2]sopBSPI-1F:GGACCGCCCAGCAACAAAACAAGAAGAAGR:AGTGATGCCCGTTATGCGTCAGTGTATT220[2]sifASPI-2F:TTTGCCGAACGCGCCCCCACACGR:GTTGCCTTTTCTTGCGCTTTCCACCCATCT449[2]sscASPI-2F:ATGAAAAAAGACCCGACCTAR:TTAGCTCCTGTCAGAAAGTT474[3]sseESPI-2F:ATGGTGCAAGAAATAGAGCAR:TTAAAAACGTCGCTGGATAA417[3]spvB毒力质粒F:GGTTACTGCATGACAGTAACGGCR:CGCAAAGCTTGTTCAGTATCGG561[2]pefA毒力质粒F:GCGCCGCTCAGCCGAACCAGR:CAGCAGAAGCCCAGGAAACAGTG157[2]spvR毒力质粒F:AGGAAATCGGACCTACGGR:TAACATCGCCAGCCCTTG473[3]spvC毒力质粒F:AAGGTCGTTCAACAAGCCR:CATTTCACCACCATCACG252[3]
1.3数据处理 应用EXCEL2007软件进行数据整理,利用SPSS18.0软件包进行统计分析,检验水准α=0.05。
2 结 果
2.1菌株复核鉴定结果 所有239株待检菌株经VITEK 2 Compact GN鉴定卡复核鉴定,均提示为沙门菌。经血清复核鉴定,结果为:肠炎沙门菌91株、鼠伤寒沙门菌58株,这2种血清型沙门菌占了62.3%(149/239),其余详见表2。
2.2各种毒力基因片段检出情况 239株沙门菌均检出invA和sopB,检出率为100%(239/239);237株检出sseE,检出率为99.2%(237/239);227株检出sscA,检出率为95.0%(227/239);204株检出sifA,检出率为85.4%(204/239);170株检出spvC,检出率为71.1%(170/239);pefA、spvB、spvR等3种基因均有111株菌检出,检出率为46.4%(111/239)。
2.3毒力基因片段阳性检出数及结果模式 239株沙门菌中,共检出毒力基因1 649个,平均每菌株检出毒力基因6.9种。其中,95株检出全部9种毒力基因,占39.8%(95/239);67株检出5种毒力基因,占28.0%(67/239);44株检出6种毒力基因,占18.4%(44/239);此3类合计为86.2%。根据不同毒力基因片段的阳性检出结果不同,可得到不同的阳性检出结果模式,而不同血清型沙门菌主要检出的毒力基因片段的模式也不同。检测菌株数较多的肠炎沙门菌、鼠伤寒沙门菌、斯坦利沙门菌、都柏林沙门菌的毒力基因阳性检出结果共有13种模式(详见表3)。其中,肠炎沙门菌以模式1(即检出全部9种毒力基因)为主,有76株,占83.5%(76/91);鼠伤寒沙门菌以模式12、模式10(invA、sopB、sseE、sscA、sifA等5种毒力基因阳性,spvB、spvR、pefA等3种毒力基因阴性,spvC阴性或阳性)为主,共45株,占77.6%(45/58);斯坦利沙门菌也是以模式10、模式12为主,共8株,占66.7%(8/12);都柏林沙门菌以模式1为主,有3株,占37.5%(3/8)。可见,在这4种常见血清型沙门菌中,模式1、模式10、模式12是比较主要的毒力基因阳性结果模式。
表2 龙岩市不同血清型沙门菌毒力基因检出情况
Tab.2 Virulence gene detection of different serotypes ofSalmonellain Longyan City

沙门菌血清型名称菌株数invAsopBsseEsscAsifAspvCspvBspvRpefA毒力基因数阳性菌株数(%)阳性菌株数(%)阳性菌株数(%)阳性菌株数(%)阳性菌株数(%)阳性菌株数(%)阳性菌株数(%)阳性菌株数(%)阳性菌株数(%)总数每菌株平均数肠炎91911009110091100911008087.9 8694.5 8795.6 8795.6 8087.9 7848.6 鼠伤寒58581005810058100581005798.3 3255.2 1017.2 1119.0 1220.7 3546.1 斯坦利12121001210012100121001083.3 975.0 18.3 18.3 216.7 715.9 都柏林88100810081008100787.5 675.0 675.0 562.5 450.0 607.5 纽波特5510051005100510051005100000000306.0 乙型副伤寒551005100510051005100360.0 000000285.6 德尔卑44100410041004100375.0 125.0 000000205.0 里森331003100310031003100133.3 000000165.3 伤寒221002100210021002100210021002100150.0 178.5 猪霍乱2210021002100210021002100000000126.0 阿贡纳111001100110011001100110000000066.0 海德尔堡111001100110011001100110000000066.0 鸡111001100110011001100110011001100110099.0 科特布斯1110011001100110011000000000055.0 鲁齐齐1110011001100110011000000000055.0 伦敦111001100110011001100110000000066.0 明斯特1110011001100110011000000000055.0 山夫登堡1110011001100110000.0 0000000044.0 圣保罗1110011001100110011000000000055.0 印第安纳1110011001100110011000000000055.0 未分型3939100391003794.92769.22153.8 1948.7410.3410.31128.22015.2合计23923910023910023799.222795.020485.417071.111146.411146.411146.416496.9
表3 肠炎沙门菌、鼠伤寒沙门菌、斯坦利沙门菌、都柏林沙门菌各毒力基因阳性检出结果模式
Tab.3 Virulence gene positive detection patterns ofS.enteritidis,S.typhimurium,S.stanleyandS.dublin

模式invAsopBsseEsscAsifAspvCspvBspvRpefA肠炎沙门菌鼠伤寒沙门菌斯坦利沙门菌都柏林沙门菌1+++++++++7610132+++++-+++10003++++-++++20004++++++-++01005+++++++-+00016++++++++-00027++++--+++10008++++-+++-70009++++++--+011010++++++---0205011++++-+---102012+++++----3253113++++-----0101
注:“+”表示阳性检出,“-”表示未检出。

2.5不同年代沙门菌毒力基因阳性检出菌株数比较 由于invA、sopB等2种毒力基因的检出率均为100%,所以在所有年代都无差异;sseE、spvC在不同年代间也没有差异,显示了较好的稳定性;而sifA、spvC、spvB、spvR、pefA等5种毒力基因在不同年代沙门菌中最少检出数差异具有统计学意义,表明龙岩沙门菌携带这5种毒力基因的菌株数量随着时间的进程而增加,详见表4。
表4 龙岩市不同年代沙门菌9种毒力基因阳性检出菌株数比较
Tab.4 Comparison of nine virulence gene positive strains ofSalmonellain Longyan in different years

年代检测菌株数invAsopBsseEsscAsifAspvCspvBspvRpefA1991-20001111111111100082001-2010424242414136271212132011-2017186186186185185167133999990趋势卡方值(χ2)///1.5095.9431.993.0718.2618.267.39P值///=0.5<0.001<0.0010.2<0.001<0.001=0.025
2.6人体样本和食品样本分离株毒力基因阳性检出数比较 人体样本分离的沙门菌以检出9种、5种、6种毒力基因的菌株为主,共157份,占87.8%,食品样本分离的沙门菌以检出9种、5种毒力基因的菌株为主,共45株,占75%,均明显高于携带其它毒力基因种数的菌株。但两类不同来源沙门菌毒力基因阳性检出菌株数差异无统计学意义(χ2=0.7,P=0.998)。见表5。
表5 不同来源沙门菌不同种类数毒力基因阳性检出菌株数比较
Tab.5 Comparison of virulence gene positive strains ofSalmonellafrom different sources
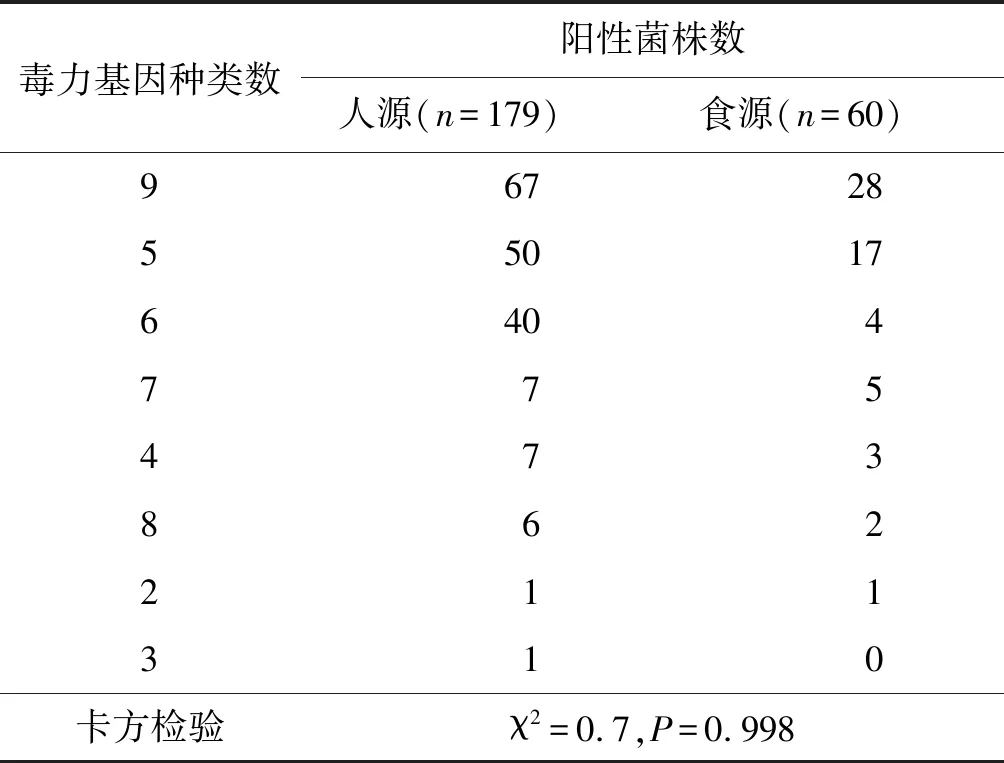
毒力基因种类数阳性菌株数人源(n=179)食源(n=60)96728550176404775473862211310卡方检验χ2=0.7,P=0.998
3 讨 论
沙门菌是与人类关系最密切的致病菌,可引起食物中毒,以及胃肠炎、菌血症或败血症、肠热症等多种疾病。本研究结果表明,龙岩市沙门菌感染以肠炎沙门菌与鼠伤寒沙门菌为主,共占62.3%(149/239)。毒力是沙门菌致病的根本原因,是由分布于其中的许多毒力基因相互作用而产生的。沙门菌毒力基因主要由染色体上的毒力岛(Salmonella pathogenicity island,SPI)编码,少数由毒力质粒(plasmid)编码[4]。
目前已发现的SPI有SPI-1、SPI-2、SPI-3、SPI-4、SPI-5等20多个[5]。其中SPI1遗传性稳定,是沙门菌入侵宿主非吞噬细胞所必需的,在侵袭巨噬细胞和肠上皮细胞过程中发挥重要作用,几乎所有沙门菌血清型都具有[6]。SPI2主要控制沙门菌在吞噬细胞和上皮细胞内进行复制,使其逃避巨噬细胞辅酶依赖的杀伤作用。SPI2编码与系统感染有关的Ⅲ型分泌系统,主要有sse、ssc、ssa和ssr等基因。其中,sse编码分泌系统效应蛋白,ssc编码分子伴侣,ssa编码Ⅲ型分泌系统成分,ssr编码分泌系统调节子[3]。所以,本研究重点选择SPI1中的invA与sopB毒力基因,SPI2中的sifA、sseE、sscA毒力基因进行研究。
invA是沙门菌属特异性基因[7],是主要毒力因子,决定沙门菌的侵袭力,与其致病性密切相关[8]。sopB基因编码参与肠黏膜液体分泌和炎症反应的相关蛋白。invA与sopB这2种基因都是SPI标志性基因[9],在本研究中,其检出率均为100%,与杨劲松(福建省临床肠炎沙门菌)[2]、夏宾雁(四川山羊源沙门菌)[10]、孙璐(山东肉鸡屠宰生产链中沙门菌)[11]研究结果一致,与彭斌[12]从新疆零售牛肉中沙门菌的研究结果相比,invA的检测结果一致,而sopB检出却相差很大(7.1%),这是否与沙门菌分离的地域或物种有关尚待研究。SPI2中的3种毒力基因检出率都在85%以上。曹恬雪等研究表明携带SPI1+SPI2与沙门菌的致病性呈正相关[13],研究表明龙岩市沙门菌的毒力较强。
在沙门菌的毒力质粒中,spv(Salmonella plasmid virulence genes)是最重要的毒力质粒,包括spvA、spvB、spvC、spvD、spvR、pefA等基因,有助于提升沙门菌在宿主细胞内的粘附能力、存活时间及繁殖能力等,从而提升其在宿主细胞内的生存能力[14]。spv表现为细胞毒性,与肠系膜淋巴结、脾、肝等肠道外组织感染有关[3]。pefA基因是菌毛操纵子基因,在感染阶段发挥重要作用,通过伴侣诱导的装配途径调节细菌对肠上皮细胞的粘附[15]。pefA、spvC基因是沙门菌毒力质粒上特有的标志性基因[1,9]。
毒力质粒的携带与特定的血清型有关,目前已报道的携带毒力质粒的沙门菌血清型有11种,包括鼠伤寒沙门菌、猪霍乱沙门菌、肠炎沙门菌、都柏林沙门菌、丙型副伤寒沙门菌、鸡沙门菌、布利丹沙门菌、御成门沙门菌、阿贡纳沙门菌、S.sendai、S.abortusovis[10]。本研究中,不仅从肠炎沙门菌、鼠伤寒沙门菌、都柏林沙门菌、猪霍乱沙门菌、阿贡纳沙门菌、鸡沙门菌中检出spvC,还从伦敦沙门菌、斯坦利沙门菌、纽波特沙门菌、乙型副伤寒沙门菌、德尔卑沙门菌、里森沙门菌、伤寒沙门菌、海德尔堡沙门菌等8种血清型沙门菌中检出spvC,但仅从肠炎沙门菌、鼠伤寒沙门菌、都柏林沙门菌、鸡沙门菌、斯坦利沙门菌、伤寒沙门菌中检出pefA。这可能与样品来源及地域差异有关,值得进一步研究。
本研究中,肠炎沙门菌4种质粒毒力基因的检出率均在87%以上,与鼠伤寒沙门菌的阳性检出率差异有统计学意义。其它血清型沙门菌由于菌株数较少,有待收集到更多菌株后再作进一步比较分析。
本研究中沙门菌spvC检出率为71.1%,其中肠炎沙门菌spvC阳性检出率更是高达94.5%:分离自人体样本的63株肠炎沙门菌58株检出spvC,阳性率为92.1%(58/63),分离自食品样本的28株肠炎沙门菌均检出spvC,阳性率100%,没有统计学差异(χ2=1.07,P=0.3)。龙岩市spvC的检出率高于庄孝飞2008-2012年上海(食源性+医源性)肠炎沙门菌分离株中spvC的检出率(78.9%)[1]、远高于夏宾雁山羊源沙门菌spvC的阳性检出率(47.6%)[10]和张芬2006-2013年间于宁波地区沙门菌中spvC的检出率(21.9%)[16],更有甚者,彭斌在新疆零售牛肉中分离的沙门菌中未检出spvC[12]。表明龙岩市沙门菌特别是肠炎沙门菌spvC毒力基因携带率高于全国各地的水平,同时由于龙岩市沙门菌spvC毒力基因在不同年代间相对稳定,那么这种差异可能与各种血清型沙门菌构成不同、地域不同、来源物种不同有关,有待进一步研究分析。
本研究结果还显示,龙岩市沙门菌除invA、sopB、sseE、spvC等4种毒力基因阳性检出数量随着时间的进程相对稳定外,其余5种毒力基因特别是质粒毒力基因的阳性检出数随时间进程呈现显著性增加,表明龙岩市沙门菌质粒毒力基因有累积增强趋势,这一方面主要与质粒本身的特性有关:质粒存在于细菌细胞浆中,独立于染色体而存在,是细菌重要的遗传物质,可提高细菌选择有利生存条件的能力,使细菌在特殊的环境下生存和生长,并且通过在种内、种间甚至属间进行DNA交换转移使其生存能力不断因进化而增强,并同时实现毒力基因的累积增强;另一方面也可能这些毒力基因是以转座子等其它因子形式存在于菌体内的,进而增强了菌株的毒力。
根据刘芳萍[17]、黄瑞[18]等的研究结果,携带毒力基因较多的菌株致病性也较强,特别是携带spvC基因的菌株大多数表现为强致病性。本次研究的9种毒力基因在龙岩市沙门菌的检出率均高于45%,平均每菌株检出毒力基因数为6.9种,检出毒力基因达5种以上的菌株数共226株,占94.56%。所有菌株均检出invA、sopB2种SPI1毒力基因,绝大部分菌株检出SPI2毒力基因中的sseE、sscA、sifA,超过70%菌株检出spvC,spvB、spvR、pefA阳性检出率均为46.4%。其中肠炎沙门菌以检出全部9种毒力基因为主,占83.5%(76/91);鼠伤寒沙门菌以invA、sopB、sseE、sscA、sifA阳性,spvB、spvR、pefA阴性结果多见,占77.6%(45/58)。结果表明龙岩市沙门菌携带较多的毒力基因,臻病性较强。本研究结果还显示食源性与人源性分离株携带毒力基因无显著性差异,表明龙岩市食源性与人源性沙门菌交叉污染/感染特别严重,应同步加强人、禽、畜沙门菌病的监测与管理。
利益冲突:无
