檫木家系生长、叶色遗传变异及优良家系选择
2019-09-17张伟民包志彬汪秀玲郑文华
张伟民 包志彬 汪秀玲 郑文华
檫木家系生长、叶色遗传变异及优良家系选择
张伟民1包志彬2汪秀玲1郑文华1
(1 浙江省开化县林场;2 浙江省开化县杨林镇林业站 324300)
以檫木半同胞家系为材料,揭示家系苗期生长、叶片颜色和叶片色素等主要性状变异规律,并利用层次分析方法对其进行综合评价。研究结果表明,檫木生长性状中的叶量、苗高、基径和分枝数,叶片颜色中的叶色a*值、叶色b*值和叶色L值在家系间具有显著的差异;以上7个性状受中度以上的遗传力控制。叶片花色素苷含量、类胡萝卜素含量和叶绿素含量在家系间差异不显著,并且具有较低的广义遗传力,说明这3个性状受环境影响较大。通过层次分析法对檫木家系进行综合评价,筛选出苗期表现比较优异的家系。檫木家系间生长和叶色性状遗传变异丰富,遗传改良潜力较大,并筛选出来自浙江和安徽的3个优良家系。
檫木;家系;遗传变异;层次分析法;选择
檫木((Hemsl.)Hemsl)是樟科(Lauraceae)檫木属植物,为高大落叶乔木;其天然分布广泛,长江流域以南各地均有分布,主要生长于海拔150-1900米疏林或密林中,是一种适应性很强的乡土树种[1]。檫木树干圆满通直,叶片形状多变,初春黄花报春,夏季绿冠如伞,秋季红叶飞扬,冬季老干挺拔,是江南生态林彩化的重要彩叶树种。另外,檫木木材浅黄色,质地良好,纹理美观,是优良的用材树种,具有很高的观赏和经济价值[2,3]。国内檫木相关研究起步于上世纪七十年代,全国开展了大规模的檫木地理种源试验[4]。檫木不同种源间苗期和幼林期在生长、抗寒性、材积等性状差异显著,并筛选出了苗期和幼林期适宜本地发展的优良种源[5-7]。孙鸿有等[8]研究认为优良檫木的选择应该在优良种源内进行家系选择。檫木主要的天然群落类型是针阔混交林,檫木能有效促进混交树种如杉木[9,10]、国外松[11]和毛竹[12]的生长以及根系营养元素吸收,因而林分生态效益突出。
党的十八届五中全会上明确了绿色发展理念,建设“美丽中国”的奋斗目标。彩叶树种由于兼具了经济价值、生态价值和观赏价值,日益受到重视[13],枫香[14]、榉树[15]、无患子[16]等一批彩叶树种陆续开展了良种选育工作。彩叶树种其生长、叶色、叶片色素含量等性状的遗传变异规律和遗传稳定性的理论研究是选育优良秋色叶树种的前提[17]。何贵平等[18]对枫香地理种源幼林的树高和地径研究发现,树高有较高的广义遗传力,与种源地理经度和纬度间均呈显著负相关,而地径则未表现出显著负相关;并利用独立选择法初选3个幼林期较速生的种源。窦全琴等[19]对榉树43个优树子代苗期和幼林期生长测定结果表明,优树子代间苗高、地径、分枝数等生长性状均有极显著差异,其变异受中等以上遗传力控制,具有很大的遗传改良潜力。榉树单株的叶色每年能保持一致,说明其叶色性状已经稳定[20]。黄利斌等研究发现榉树种子千粒重、发芽势、场圃发芽率、苗高、地径、Ⅰ级侧枝数和秋季叶色变化等性状在家系间均存在显著差异,且不同叶色的植株比例与母树秋色叶也存在相关性[21]。
前期有关彩叶树种遗传改良主要针对生长性状进行选育,较少考虑其叶片颜色、叶片色素含量等性状指标。所以本文以檫木半同胞家系为材料,分析其苗期生长、叶色、叶片色素含量等性状遗传变异规律,并结合层次分析法[22]对檫木家系生长、叶色、叶片色素含量等性状进行综合评价,以期筛选生长和观赏兼优的家系。
1 试验材料与方法
1.1 试验材料
2013年6月底到7月中下旬,采集了湖南、湖北、浙江、安徽、江西、福建6个省区20个檫木优良单株的种子(表1)。2014年3月初在圃地进行育苗,随机区组试验,每小区栽植10株,6次重复。育苗地位于东经119°27′E、北纬29°41′N,属于典型的亚热带季风气候,年平均气温17.80 ℃,年平均降水量1486.80 mm,年日照时数1948.50 h。
1.2 试验方法
2014年11月中旬对圃地的檫木苗进行调查,调查的指标主要包括叶量(檫木叶片的数量)、苗高、基径、分枝数、叶色L值、叶色a*值、叶色b*值、叶绿素含量、类胡萝卜素含量和花色素苷含量等。叶量和分枝数采用计数的方法进行调查,苗高主要用皮尺测量,基径用游标卡尺测定。檫木叶片颜色值(L、a*、b*)的测量采用分光测色计CM-700d(日本,柯尼卡)进行测定。其中,L表示叶片的色彩亮度,a*表示从绿色到红色的色相值,b*表示从蓝色到黄色的色相值[23]。采用紫外分光光度计(UV-1800A,上海美析)对叶绿素含量、类胡萝卜素含量和花色素苷含量进行测定。
表 1 檫木20个家系的地理位置
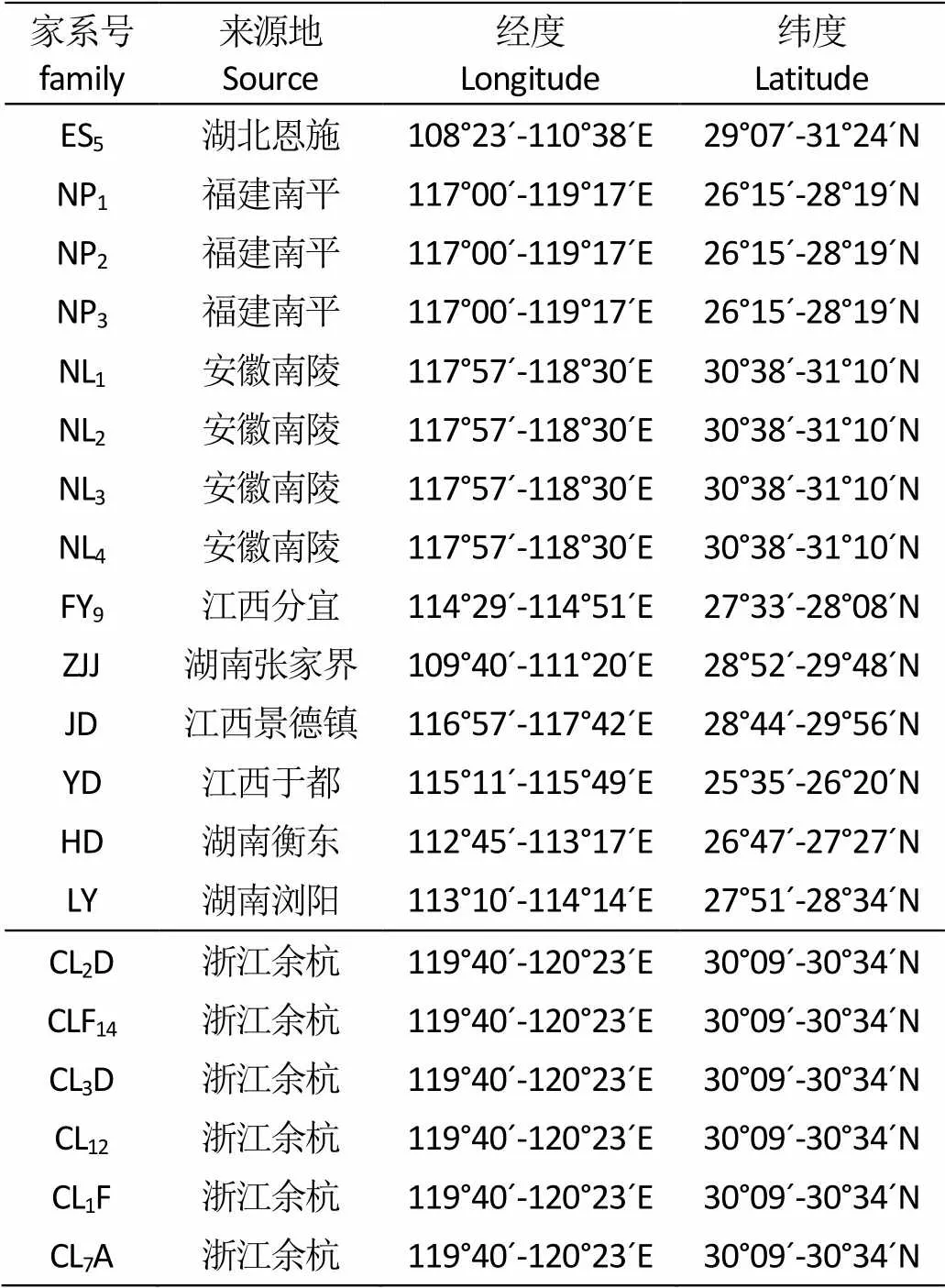
Tab.1 Geographic location for twenty families of Sassafras tsumu
1.3 层次分析法
1.3.1建立层次结构模型
将檫木秋色叶优良观赏品系的综合评价视为复杂的系统问题,选出最优观赏家系是其终极目标(目标层A),围绕终极目标所构建的指标评价体系既要关注檫木的生长性状,更要关注它的叶色和色素含量,具体指标见图1。
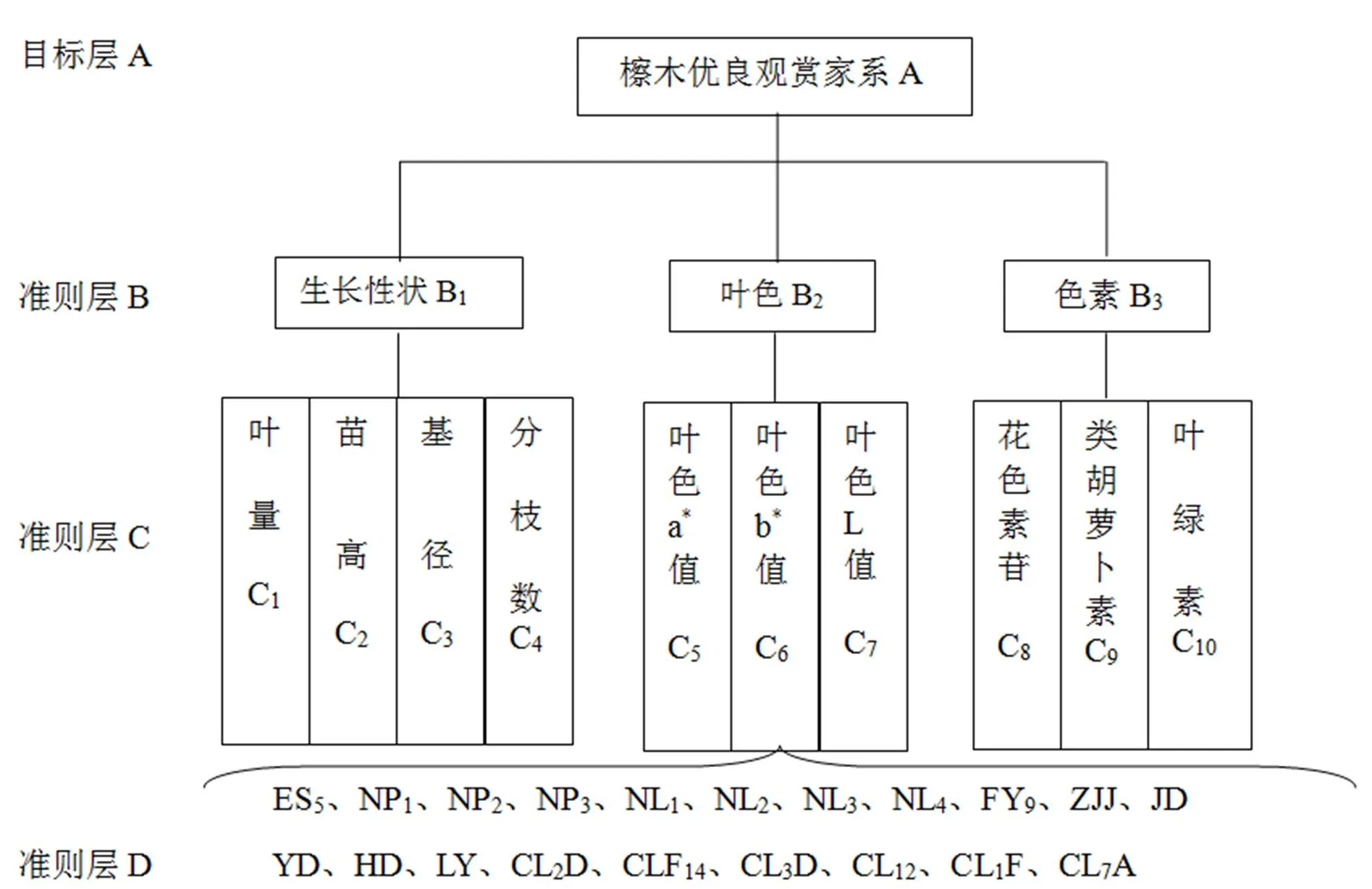
图1 檫木秋色叶优良观赏家系选择指标评价体系
1.3.2构造判断矩阵
构造判断矩阵一般采用1~9及其倒数的标度方法,即把各评价因素从劣到优均划分为9个等级,同一层的因素两两比较得出相对于上一层因素的相对重要值。两因素相比较,分别用1、3、5、7、9表示相互之间的相对重要性。2、4、6、8表示介于1、3、5、7、9相邻判断的中间值。
1.3.3层次单排序及其一致性检验
由判断矩阵计算最大特征根,然后得出特征根对应的特征向量;下一层因素对于上一层相应因素的相对重要性排序权值用规一化后的特征向量值表示。其中和的计算公式如下:


表2 A-B判断矩阵
Tab 2 A-B comparison matrix

注:W1表示B1、B2、B3相对于A的权重值,CR1表示A-B的随机一致性比率。
表3 B1-C判断矩阵
Tab 3 B1-C comparison matrix

注:W2表示C1、C2、C3、C4相对于B1的权重值,CR2表示B1-C的随机一致性比率。
表4 B2-C判断矩阵
Tab 4 B2-C comparison matrix

注:W3表示C5、C6、C7相对于B2的权重值,CR3表示B2-C的随机一致性比率。
表5 B3-C判断矩阵
Tab 5 B3-C comparison matrix
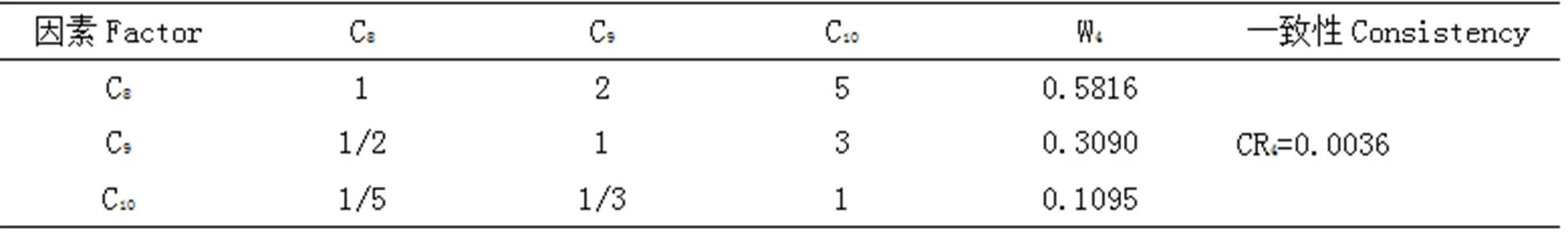
注:W4表示C8、C9、C10相对于B3的权重值,CR4表示B3-C的随机一致性比率。
1.3.4 层次总排序及一致性检验
准则层C所有因素相对于目标层A的重要性排序以层次总排序表示。因为CR=0.000 <0.10,所以认为层次总排序的结果有满意的一致性。通常当准则层D的因素数量超过9个时,一般采用绝对评价的数量学方法计算其综合数量评价值[24]。因为进行檫木秋色叶优良观赏品系的综合评价时,准则层D的家系数量为20个,已经远远超过9,所以采用绝对评价的数量学方法对檫木秋色叶优良观赏品析进行选择。檫木秋色叶优良观赏品系综合数量评价值()的计算公式如下:
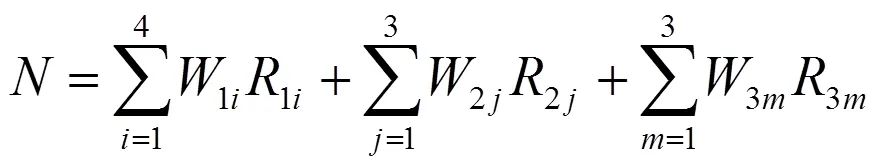
1.4 数据分析
利用DPS11.5软件对檫木秋色叶优良观赏品系的各性状进行层次分析和方差分析,广义遗传力用续九如[25]的方法进行估算。相关图表用Sigmaplot 10.0和Microsoft Excel 2007软件绘制。
2 结果与分析
2.1 檫木子代生长性状的变异分析
通过对檫木苗20个家系生长性状(叶量、苗高、基经和分枝数)的统计分析可知(表6)。檫木20个家系中,浙江余杭CLF14家系的平均叶量值最大、为22.30,平均叶量最小值是湖北恩施ES5家系、为4.00,最大值是最小值的5.6倍;浙江余杭CL1F家系的平均苗高值最大、为108.50 cm,平均苗高最小值是湖南衡东HD家系、为41.07 cm,最大值是最小值的2.6倍;浙江余杭CL1F家系的平均基径值最大、为9.48 cm,平均基径最小值是江西于都YD家系的5.65 cm。分枝数平均最大值是浙江余杭CLF14家系、为3.50,最小值是江西于都YD家系的1.10,最大值是最小值的3.2倍。
由表7可知,叶量、苗高、基径和分枝数4个生长性状指标在家系间具有极显著的差异。通过对4个生长性状的广义遗传力的分析可知(表7),叶量、苗高、基径和分枝数的广义遗传力分别为0.73、0.95、0.84和0.65。说明参试家系的苗高、基径、叶量和分枝数4个生长性状受中度以上的遗传力控制。
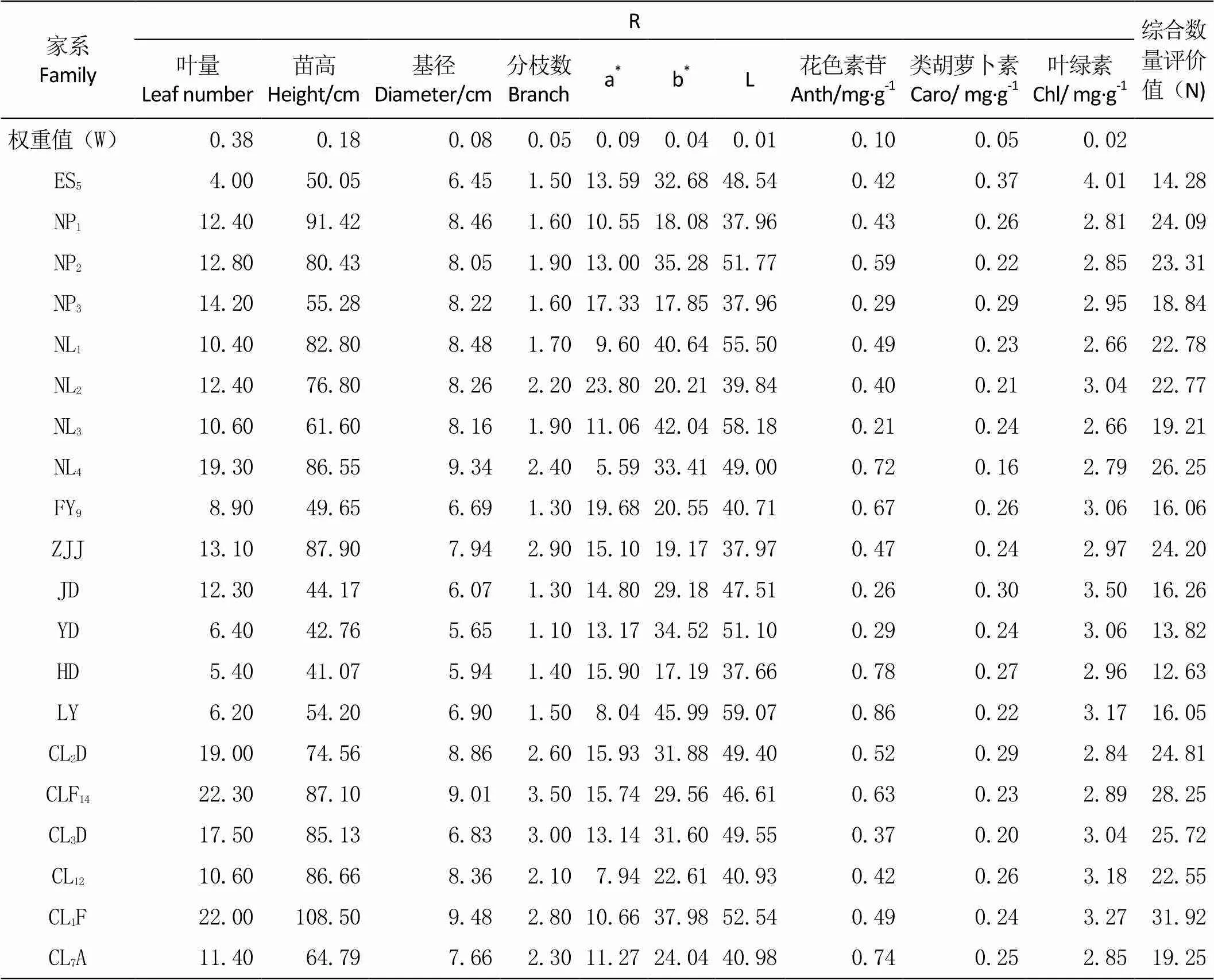
表6 檫木20个家系主要性状及综合数量评价
注:W表示准则层C相对于目标层A的权重值。
2.2 檫木子代叶色性状的变异分析
由表6可知,平均叶色a*值最大值是安徽南陵NL2家系的23.80,最小值是安徽南陵NL4家系的5.59,最大值是最小值的4.3倍;湖南浏阳LY家系的平均叶色b*值最大、为45.99,湖南衡东HD家系的平均叶色b*值最小、为17.19。平均叶色L值最大值是湖南浏阳LY家系的59.07,最小值是湖南衡东HD家系的37.66。叶色值的方差分析表明(表7),叶色a*值、叶色b*值和叶色L值在家系间存在极显著的差异。叶色a*值的广义遗传力为0.74,叶色b*值和叶色L值的广义遗传力均为0.85,表明叶色受遗传因子的影响比较大。
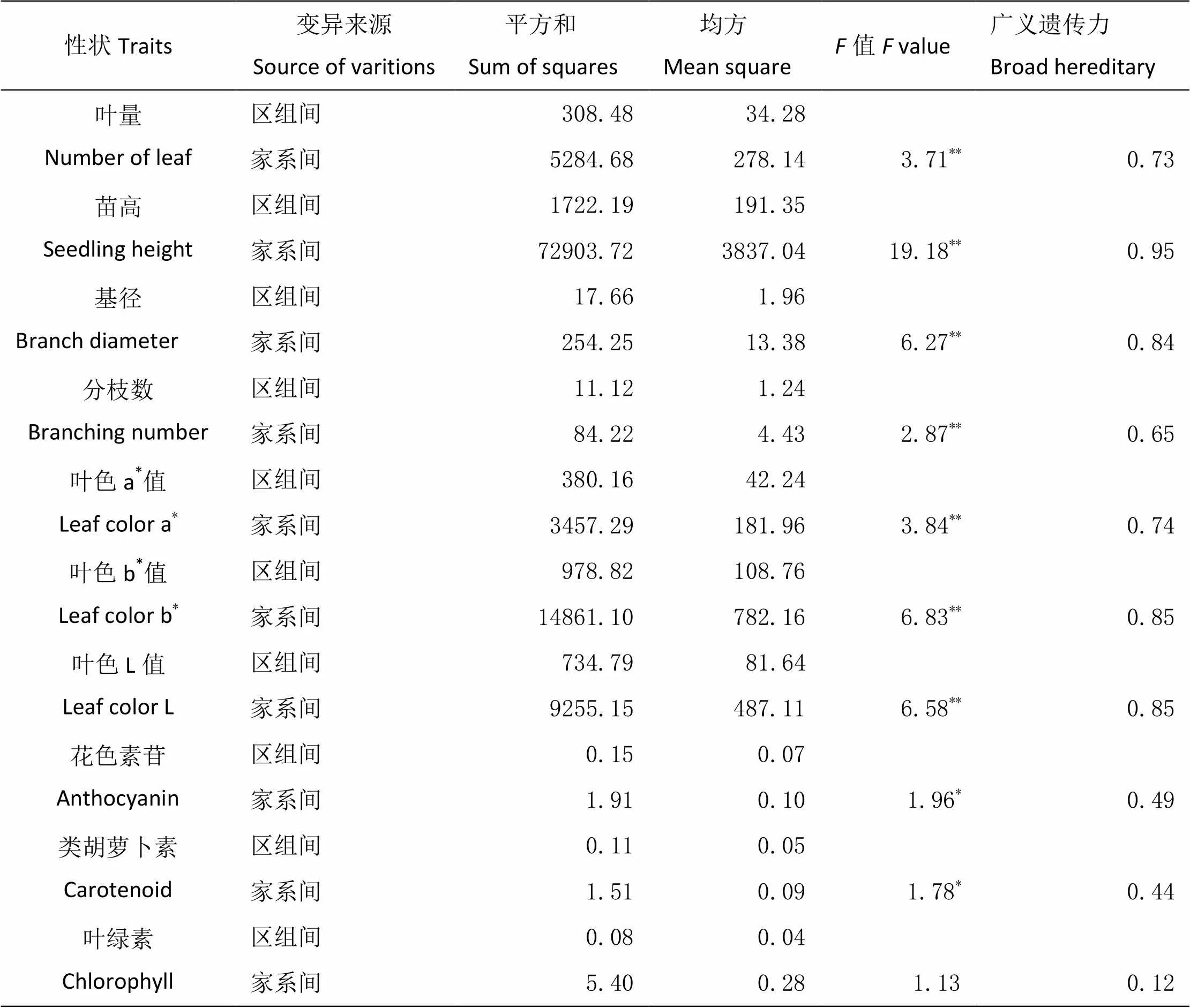
表7 檫木20个家系主要性状的方差分析
注:**表示显著性概率为0.01,*表示显著性概率为0.05。
2.3 檫木子代色素含量的变异分析
色素含量是植物的生理性状,受环境和遗传因子的双重影响。对檫木20个家系色素含量的分析可知(表6),湖南浏阳LY家系的平均花色素苷含量最大、为0.86 mg·g-1,安徽南陵NL3家系的平均花色素苷含量最小、为0.21 mg·g-1,最大值是最小值的4.1倍。平均类胡萝卜素最大值是湖北恩施ES5家系的0.37 mg·g-1,最小是安徽南陵NL4家系的0.16 mg·g-1。安徽南陵ES5家系的平均叶绿素含量最大、为4.01 mg·g-1,安徽南陵NL1家系的平均叶绿素含量最小为、2.66 mg·g-1。由表7可知,花色素苷含量和类胡萝卜素含量在家系间存在显著差异,然而叶绿素含量在檫木家系间不具有显著性差异。花色素苷含量、类胡萝卜素含量和叶绿素含量的广义遗传力分别为0.49、0.44和0.12。由此说明,檫木叶片的色素含量广义遗传力较低,色素含量受环境因子的影响较大,尤其是叶绿素含量具有极低的广义遗传力,说明立地环境因子影响着檫木叶片内叶绿素的合成。
2.4 檫木秋色叶优良观赏品系选择
根据表6的权重值(W)可知,在生长性状、叶色值和色素含量共10个指标中,叶量的权重值最大为0.38,苗高的权重值次之为0.18,叶色L值的权重值最小为0.01。说明在檫木秋色叶优良观赏品系的选择中,叶量和苗高占有很重要的分量。由20个檫木家系的综合数量评价值可知(表6),檫木秋色叶优良观赏品系从优到劣的排序为:CL1F>CLF14>NL4>CL3D>CL2D>ZJJ>NP1>NP2>NL1>NL2>CL12>CL7A>NL3>NP3>JD>FY9>LY>ES5>YD>HD。从排序可知,来自浙江余杭CL1F和CLF14家系和安徽NL4家系等在浙江地区具有良好的表现;而来自湖北、江西和湖南的ES5、YD和HD等家系在浙江地区表现相对较差。
3 结论与讨论
3.1 檫木苗期生长、叶片变异规律
上世纪八、九十年代国内学者就开始了檫木种源、类型和家系三个层面的选育,为檫木后期的研究奠定了坚实的基础。在长期的檫木选优中发现,檫木选优应该在优良种源内进行家系选择[5,6]。所以,本文利用所收集的檫木优良家系材料,从其苗期生长表现和秋色叶颜色、叶片色素含量等性状指标出发,探讨其生长和叶色等性状遗传变异规律,并通过层次分析法对檫木秋色叶优良观赏品系进行选择。研究发现,檫木生长性状中的叶量、苗高、基径和分枝数,叶色中的叶色a*值、叶色b*值和叶色L值在家系间具有显著的差异,此7个性状受中度以上的遗传控制,说明这7个性状具有很大的异常改良潜力。孙鸿有等[8]对檫木家系的研究也发现,树高和胸径在家系间存在显著的差异。张谦等[26]对同为樟科的樟树进行家系选择时发现,树高、胸径和分枝数在家系水平上存在显著差异,家系遗传力在0.56-0.85之间,这与本究的结果基本一致。但本研究发现檫木叶片的花色素苷含量、类胡萝卜素含量和叶绿素含量在家系间差异不显著,并且具有较低的广义遗传力,说明这3个性状受环境影响较大。
3.2 檫木优良家系选择
层次分析法(AHP)可将复杂的问题分解为若干不同层次,并逐步分解、分析,已在农业、经济、军事等领域得到了广泛的应用[27]。蒋艾平等[28]利用层次分析法对乐东拟单性木兰进行地理种源变异的选择,筛选出适宜浙北发展的乐东拟单性木兰优良种源。本文通过层次分析法对檫木生长、叶色和生理等性状的分析表明,来自浙江和安徽的家系在浙江地区具有良好的表现,可以采集这二个省份种子在浙江地区进行推广和应用。而来自湖北、湖南和江西南部的檫木家系在浙江地区表现相对较差,这可能与光、温、水、热和土壤条件等环境因子存在较大的差异有关。孙鸿有[5]研究结果也表明浙江种源苗期适应性和生长等性状综合表现也较好,适宜浙江地区发展,所以推荐使用该省种源的种子。
[1] 中国科学院中国植物志编辑委员会. 中国植物志第30卷[M]. 北京:科学出版社,1959:147.
[2] 成俊卿. 木材学[M]. 北京: 中国林业出版社,1985:10-23.
[3] 蒋艾平,姜景民,刘军. 檫木叶片性状沿海拔梯度的响应特征[J]. 生态学杂志,2016,35( 6) : 1467-1474.
[4] 罗勤初,肖国华. 檫木地理种源试验幼林阶段初报[J]. 湖南林业科技,1986,3(2): 25-28.
[5] 孙鸿有,徐明浪, 万寿余. 檫树不同地理种源苗期变异的初步研究[J]. 浙江林学院学报, 1985, 2(2):1-6.
[6] 孙鸿有,何允铨,苏建国. 种子产地对擦树幼林变异影响的研究[J]. 1989, 42(4): 16-21.
[7] 肖国华, 罗勤初, 罗中甫. 湖南檫树种源选择的研究[J]. 湖南林业科技, 1992,(4): 10-15.
[8] 孙鸿有,陈赛娟,宋小友. 檫树种源、类型、家系三水平选择的研究[J]. 浙江农林大学学报,1993,(7):1-2.
[9] 方志伟. 杉檫混交根系效应的研究[J]. 林业科学, 2001, 37(1):154-157.
[10]封磊, 洪伟, 吴承祯,等. 不同类型杉木人工混交林乔木层结构及动态分析[J]. 植物资源与环境学报, 2008, 17(1): 48-53.
[11]吕其根, 石连贵. 檫木、国外松混交林对檫木生长的影响[J]. 江苏林业科技, 2000, 27(2): 48-49.
[12]刘桂华, 李宏开. 檫树毛竹混交林中毛竹鞭根的研究[J]. 应用生态学报, 2002, 13(4): 285-389.
[13]侯元凯,蓝正勇,张根梅. 我国杨柳科彩叶树种种质资源及应用[J]. 世界林业研究,2017,30(2):77-81.
[14]胡文杰,王晓荣,胡兴宜,等. 枫香优良种源及家系早期选择[J]. 东北林业大学学报,2017,45(6):5-11.
[15]金晓玲,汪晓丽,刘雪梅,等. 榉树新品种‘恨天高’[J].林业科学,2016,52(4):225.
[16]刁松锋,邵文豪,陈涛,等. 无患子天然居群遗传多样性研究[J]. 林业科学研究,2016,29(2):176-182.
[17]蒋艾平. 檫木秋色叶性状变化机制研究与品系景观应用价值评价[D]. 中国林业科学研究院,2016.
[18]何贵平,陈益泰,唐雪元,等. 枫香地理种源幼林生长性状变异研究[J]. 江西农业大学学报,2005,27(4): 585-589.
[19]窦全琴,张敏,黄利斌,等. 榉树优树子代苗期及幼林期生长变异初步研究[J]. 江苏林业科技,2014, 41(3): 1-4.
[20]曹娴,罗玉兰,崔心红,等. 榉树遗传变异分析及优良单株选择[J]. 上海交通大学学报,2010,28(6): 54-56.
[21]黄利斌, 汪企明, 樊丛梅,等. 榉树半同胞家系苗期性状变异的研究[J]. 江苏林业科技,2001,28(6):1-3.
[22]吴殿廷,李东方. 层次分析法的不足及其改进的途径[J]. 北京师范大学学报(自然科学版),2004,40(2):264-268.
[23]单幼霞,邓朝军,胡文舜,等. 枇杷属植物落黄叶片颜色遗传多样性分析[J]. 园艺学报,2017,44 (4):755–767.
[24]赵焕臣. 层次分析法——一种简易的新决策方法[M]. 北京:科学出版社,1986: 77-86.
[25]续九如. 林木数量遗传学[M]. 北京: 高等教育出版社,2004:3-23.
[26]张谦,曾令海,蔡燕灵,等. 樟树自由授粉家系生长与形质性状的遗传分析[J]. 中南林业科技大学学报,2014,34(1): 1-6.
[27]王莲芬,许树柏. 层次分析法引论[M]. 北京:中国人民大学出版社,1990:150-162.
[28]蒋艾平,刘军,姜景民,等. 基于层次分析法的乐东拟单性木兰优良种源选择[J]. 林业科学研究,2015,28(1) : 50-54.
The family growth condition, heredity variation of leaf pigments and excellent family selection for
Zhang Weimin1,BaoZhibin2,WangXiuling1,ZhengWenhua1
In order to select the excellent families of, genetic variation of growth, leaf color and leaf pigment between families were investigated. Genetic variation of main traits was analyzed and evaluation of all families was conducted by using analytic hierarchy process(AHP). The results showed that there were significant differences in aspect of seven traits, including leaf number, seedling height, diameter at base height, branches, a*, b* and L among different families. This indicated that these seven traits have a huge potential for improvement. The broad heritability of these seven traits were above 0.65. This suggested that these traits were mainly controlled by genetic factors instead of environmental factors. No significant differences were found in chlorophyll, carotenoid and anthocyanin contents between different families. The broad heritability of these three traits were low, that indicated these three traits were mainling controlled by environment. Genetic variation of growth and leaf color was rich between all families. Three superior families from Zhejiang and Anhui province were selected for popularization.
; Family; Genetic variation; AHP; Selection
S722.3+3
A
1004-7743(2019)03-0012-07
2019-04-18
浙江省十三五林木育种专项课题(2016C02056-10),林业公益性行业科研专项 (201404104)。
