Helicobacter pylori virulence genes
2019-09-13AnjaterbencErikaJarcMarioPoljakMatjaHoman
Anja Šterbenc, Erika Jarc, Mario Poljak, Matjaž Homan
Abstract
Key words:Helicobacter pylori; Virulence genes; Disease association; Children; Adults;Outer membrane proteins; Bacterial toxins
INTRODUCTION
As one of the most common bacterial infections, Helicobacter pylori (H. pylori) infects approximately half of the world's population, although substantial regional variation exists[1]. The infection is usually acquired in childhood and persists lifelong in the absence of appropriate antibiotic treatment. In order to survive the harsh milieu of the human stomach, H. pylori had to adapt by possessing various virulence genes.However, the significance of these virulence genes extends beyond the pure survival needs of the bacteria, making H. pylori one of the most well-adapted human pathogens, capable of sustaining extremely efficient persistent infection. H. pylori has in fact developed mechanisms to withstand gastric acidity through the possession of urease and multiple sheathed flagella, which enable the bacteria to move toward gastric epithelial cells. H. pylori then needs to establish permanent colonization of the gastric mucosa, which is accomplished by the action of outer membrane proteins(OMPs) and adhesins, which enable adherence to the gastric epithelial cells. Finally,H. pylori possesses an arsenal of virulence genes that encode for effector proteins,which directly impair the gastric epithelium[2,3]. Although infection with H. pylori almost inevitably leads to chronic active gastritis, only approximately 10%-15% of infected individuals develop severe gastroduodenal diseases, such as peptic ulcer disease (PUD), gastric carcinoma (GC) and mucosa associated lymphoid tissue(MALT) lymphoma[4,5]. Nevertheless, the high global prevalence of H. pylori is considered an important public health issue, especially since H. pylori is classified as a class I carcinogen. More than one million (1033701) new cases of GC were estimated to occur worldwide in 2018, accounting for 6.1% of all new cancer cases, ranking GC as the fifth most common malignancy among males and females on a global scale[6].
H. pylori infection in children and adults differs in several aspects. In children, it is thought that environmental factors, such as smoking, are implicated in disease development to a far lesser degree than in adults. Whereas several factors influence the prevalence rates of H. pylori infection in children (e.g., gender, age, low socioeconomic status and family education, poor hygiene, household crowding and certain geographical regions), it has been shown that the infection is acquired in early childhood in both industrialized and non-industrialized countries[7]. The most frequent form of gastritis in children is nodular gastritis, while atrophic gastritis and intestinal metaplasia, which occur more often in adults, are relatively rarely found in children[7]. Because the degree of H. pylori colonization and repertoire of virulence genes are comparable in both children and adults, it is thought that the lower levels of gastric inflammation and lower rates of severe clinical outcome in children indicate downregulation of immune responses[8].
Over the past few decades, inclusion of proteomic and transcriptomic methods, as well as the availability of an increasing number of H. pylori partial and complete genomes, have significantly improved knowledge of the intricate gene regulatory networks of H. pylori. While the exact molecular mechanisms by which H. pylori infection induces a severe clinical outcome have not yet been clearly elucidated, they are thought to involve various elements, including host genetic and environmental factors, as well as certain bacterial virulence genes. In this review, we present the most important H. pylori virulence genes and discuss their prevalence and clinical significance in children and adults.
GENES ENCODING OUTER MEMBRANE PROTEINS
OMPs are a large group of proteins that confer durable colonization of H. pylori through specific interactions with the host receptors. It has been estimated that approximately 4% of the H. pylori genome encodes OMPs, suggesting that these proteins are of vital importance to the bacterial lifecycle[3,9]. Several OMPs have been described in detail to date, with most studies focusing on babA2, oipA, homB, and sabA genes.
babA2
To date, three allelic types of bab have been identified: babA1, babA2 and babB. The babA2 gene encodes a blood group antigen binding adhesin (BabA), a major adhesin on the outer bacterial membrane that enables binding of H. pylori to the mucosal Lewisbblood group antigens, thus facilitating colonization and determining bacterial density. Strains carrying the babA2 gene can be classified based on protein production as BabA high producers (BabA-H), which possess Lewisbbinding activity, and BabA low producers (BabA-L), which are not able to bind to Lewisbantigens, while strains carrying the babA1 gene lack BabA. Unfortunately, PCR was used in most studies evaluating the prevalence and clinical significance of babA2, although it has been shown that this method does not accurately reflect the functional status of BabA as determined by Lewisbbinding activity or immunoblotting[10,11]. Moreover, expression of BabA is generally regulated by phase variation and intragenomic recombination events between the babA gene and its highly homologous gene babB[11,12].
Adults:The prevalence of the babA2 gene varies significantly among different geographic regions, from moderate (44.0% and 44.6% in strains from Portugal and Germany, respectively) to high (70.4% and 79.7% in strains from Iran and United States, respectively) and even universal presence in strains from Japan, South Korea,Taiwan and Brazil[13]. H. pylori strains from East Asia uniformly express the BabA protein[10,14], whereas only 9.8% of Western strains were shown to lack the BabA[10].
A meta-analysis of 38 case-control studies evaluating the relationship between the presence of the babA2 and clinical outcome showed that detection of the babA2 gene significantly increases the risk of PUD [odds ratio (OR) = 2.069, 95% confidence interval (CI): 1.532-2.794], especially in the duodenal ulcer subgroup (OR = 1.588,95%CI: 1.141-2.209), with significant associations being more apparent in studies on Western isolates. Namely, the presence of the babA2 gene substantially increased PUD risk in Western populations (OR = 2.739, 95%CI: 1.860-4.032), whereas the association with PUD was only marginal in Asian populations (OR = 1.370, 95%CI: 0.941-1.994),due to the very high overall prevalence of the babA2 gene. Conversely, no significant risk correlation was observed for GC among Western (OR = 1.303, 95%CI: 0.881-1.927)or Asian populations (OR = 1.132, 95%CI: 0.763-1.680)[13]. The lack of association found in this meta-analysis could be due to significant heterogeneity among the performed studies, contradicting several reports that suggest babA2 is indeed an important virulence factor in GC development, especially when co-expressed with other virulence factors. For example, it has been shown that the “triple-positive” genotype,simultaneously containing babA2, vacA s1 and cagA, serves as a better discriminative factor for PUD and GC than the vacA s1 and cagA only genotype[15]. Moreover, a study focusing on expression of the BabA protein has shown that patients from Western countries with BabA-H and BabA-L had a 18.2- (95%CI: 1.7-198) and 33.9-fold (95%CI:2.8-411) increased risk of GC compared to those who were babA2 negative[10].Interestingly, a recent genome-wide association study on 173 European H. pylori isolates showed that, compared to strains obtained from gastritis patients, the GC phenotype was associated with certain single nucleotide polymorphisms and a specific array of genes, including the babA2 gene[16]. Although the majority of studies on isolates from East Asia have failed to find an association between the babA2 gene and disease status, a study from Taiwan highlighted the importance of the recombinant babA/B genotype, which was found to be associated with both precancerous lesions and GC[12].
Children:Data on the significance of the babA2 gene in children is less abundant. To date, nine studies have evaluated the prevalence and clinical relevance of the babA2 gene in children[17-20]. The prevalence ranged from 17.2% in Portuguese[21,22]to 84.4% in Brazilian strains[23]. Moreover, associations between the babA2 gene and clinical outcome are inconsistent[17,24,25], with only two studies[17,22]correlating the presence of the babA2 gene with a higher degree of gastric mucosal damage.
Associations with other virulence genes:The influence of the babA2 gene on clinical outcome is generally associated with cagA, vacA s1, vacA m1[17]and oipA “on”status[26,27].
Comment:Unfortunately, despite a multitude of clinical and epidemiological studies that have attempted to identify possible links between the presence of the babA2 gene and disease outcome, definite conclusions are difficult to reach, due to several factors that influence interpretation of the results. In addition to the distinct genotypic profile of Western and Asian isolates, considerable performance differences in babA2 gene detection methods[17], as well as poor correlation between the presence of the babA2 gene and actual expression and activity of the BabA2 protein[10], thus prevent simple comparisons between studies.
Outer inflammatory protein A
Outer inflammatory protein A (OipA) is encoded by the oipA gene and its expression is thought to be dependent on a slipped strand mispairing system. The proposed mechanisms by which a functional OipA (e.g., oipA “on” status) promotes severe gastric pathology include the capacity of the bacteria to attach to the gastric epithelium, followed by subsequent apoptosis of host cells, toxicity and the induction of inflammation through increased interleukin- 8 (IL-8) production[28-35].
Adults:The overall prevalence of oipA “on” status in adult patients was shown to be remarkably consistent among certain geographical regions: Approximately 100%, 80%and 60% of East Asian, Latin American and Western strains, respectively, contained oipA “on” (Table 1). Unfortunately, the clinical significance of the oipA status remains controversial, although numerous studies have investigated its relevance. It has been proposed by some authors that OipA increases the risk for PUD and GC development by disrupting the balance between apoptosis and cell proliferation during H. pylori infection, causing PUD when apoptosis is promoted and metaplasia and GC when gastric cell proliferation is increased[36-39]. A meta-analysis of PUD and GC risk, based on oipA “on/off” status, showed increased overall risk of PUD (OR = 3.97, 95%CI:2.89-5.45) and GC (OR = 2.43, 95%CI: 1.45-4.07) in individuals with oipA “on” status,while the presence of the oipA gene alone did not reflect its specific functional status,since it was not found to be associated with PUD or GC[40]. However, results from some studies contradict the findings from this meta-analysis, since no correlation between oipA and disease status or increased gastroduodenal damage was identified[27,31,32,35]. Moreover, it seems that oipA status by itself is not a useful marker for predicting the clinical outcome of H. pylori infection, especially in populations with a high prevalence of infection with virulent strains[32].
Children:In children, the frequency of the oipA “on” status tends to be somewhat lower than in adults (49.6%, 67.6% and 68.8% in children from Portugal, United States and Brazil, respectively)[22,30,41], with higher frequencies among pediatric strains from high risk populations in which the incidence of H. pylori infection and related disease is significant. Moreover, the OR for PUD risk was shown to be higher in children(OR= 7.03, 95%CI: 3.71-13.34) compared to that in adults, suggesting increased risk for PUD in children with oipA “on” status[40,42]. However, the observed differences between children and adults regarding the significance of oipA status were based on a relatively small number of strains tested and thus need to be confirmed in future studies.
Associations with other virulence genes:The oipA “on” status was found to be closely associated with cagA positivity[26,27,33,38], although it has also been linked to the presence of other H. pylori virulence genes, such as vacA s1[27,33,38], vacA m1[26,27,33], vacA m2[33]and babA2[26,27].
homB
The hom family contains four OMPs, of which homA and homB are the most studied.Strains can carry a single homA or homB gene, with one locus remaining empty, two copies of each gene (homA/homA or homB/homB), a single copy of each gene(homA/homB), or they can lack homA and homB genes, leaving both loci empty. HomB enables adherence to host gastric epithelial cells and has been shown to increase cellular IL-8 production in vitro[42]. The level of adherence and IL-8 secretion is proportional to the number of homB copies with strains that carry two copies of the homB gene, inducing more pronounced actions, leading to a higher degree of gastric mucosal damage[42].
Adults:Studies have found a relatively comparable prevalence of the homB gene in Western countries, with slightly more than half of the evaluated strains being homB positive (Table 2). However, it seems that the homB gene is more common in East Asia and West Africa than in the Middle East, where only approximately one third of strains contain homB (Table 2). In addition, the distribution, location and copy number of the homB gene seem to be dependent on geographical region, influencing potentialdifferences in disease outcome[42,43]. Whereas Western strains carry a single hom gene at locus A, East Asian strains only carry a single hom gene at locus B[42,44]. Interestingly,strains from Iran were shown to carry only one of the hom genes, homA and homB were not detected simultaneously in any of the 138 evaluated strains[45].
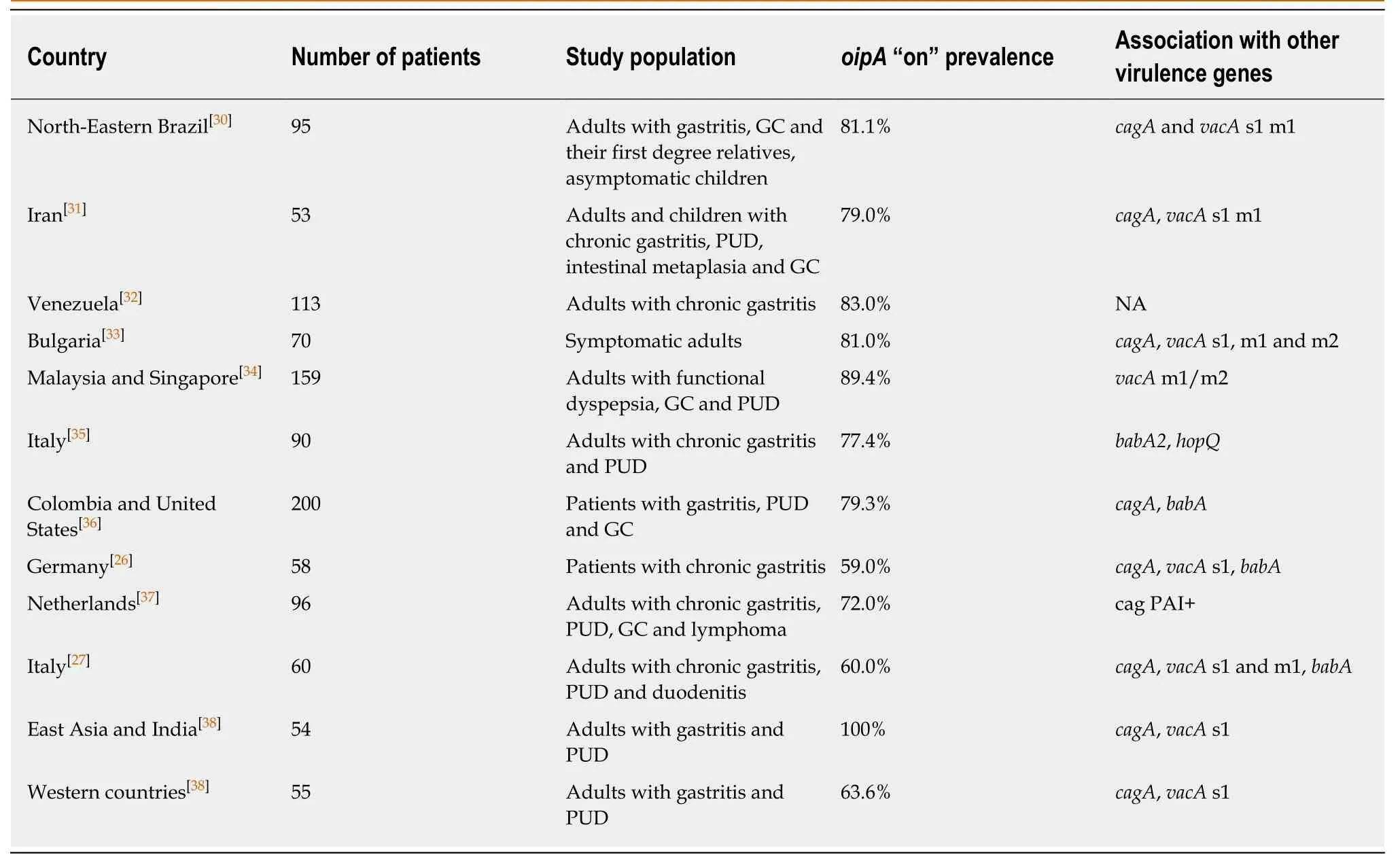
Table 1 Prevalence of oipA “on” status among isolates from various geographical regions
Whereas the two genes exhibit 90% sequence identity, they are correlated with different spectra of the disease[46-51]; homA has been associated with non-ulcer dyspepsia (NUD), whereas homB is presumed to be implicated in the development of PUD and GC, although this association is geographically dependent (Table 2).Moreover, strains carrying two copies of the homB gene were found to be most strongly correlated with PUD (OR = 4.91, 95%CI: 1.77-14.02)[42].
Children:Only three studies[22,42,50]have specifically focused on the prevalence and clinical significance of the homB gene in children. Whereas two studies from Portugal found a strong association between homB and PUD[22,42], homB was not considered to be an important individual virulence factor in Slovenian children and was only associated with a higher degree of mucosal damage when co-present with other virulence genes (i.e., cagA, vacA and babA2)[50].
sabA
In addition to Lewisbblood group antigens, sialyl-Lewisxand sialyl-Lewisaantigens are considered to be functional receptors, enabling H. pylori adherence. They are recognized by the corresponding sialic acid binding adhesin SabA, encoded by the sabA gene. In contrast to SabA, its homologue SabB does not seem to be able to bind to sialyl-Lewisxand sialyl-Lewisareceptors. Similar to oipA, the expression of SabA is regulated by phase variation, meaning only certain strains are capable of producing functional proteins[52,53]. The level of expression of SabA can rapidly adjust to the changing environment of the human stomach by switching “on” or “off”. The sialyl-Lewisxand sialyl-Lewisaantigens are otherwise rarely present in normal gastric mucosa, and only after persistent H. pylori infection induces chronic inflammation of the gastric mucosa does replacement of naturally produced Lewis antigens occur[53].Moreover, the sabA “on” status inversely correlates with the degree of gastric acid secretion, suggesting that differences in pH and/or antigen expression on atrophic mucosa can influence SabA expression[53].
Adults:In adults, sabA “on” was found in 63.2%, 49.0% and 35.5% of strains fromPortugal[42], the Netherlands[37]and Italy[35], respectively. The rates are higher in Iran,with sabA “on” being detected in 85.3% of strains[31]. Similarly, functional sabA was found to be highly prevalent in Japan, it was present in 81.5% of patients with chronic gastritis, PUD and GC[54]. Interestingly, an analysis of strains from Taiwan showed that the sabA gene was present in 80.0% (116/145) of strains, whereas only 31.0%(45/145) actually expressed SabA[14].
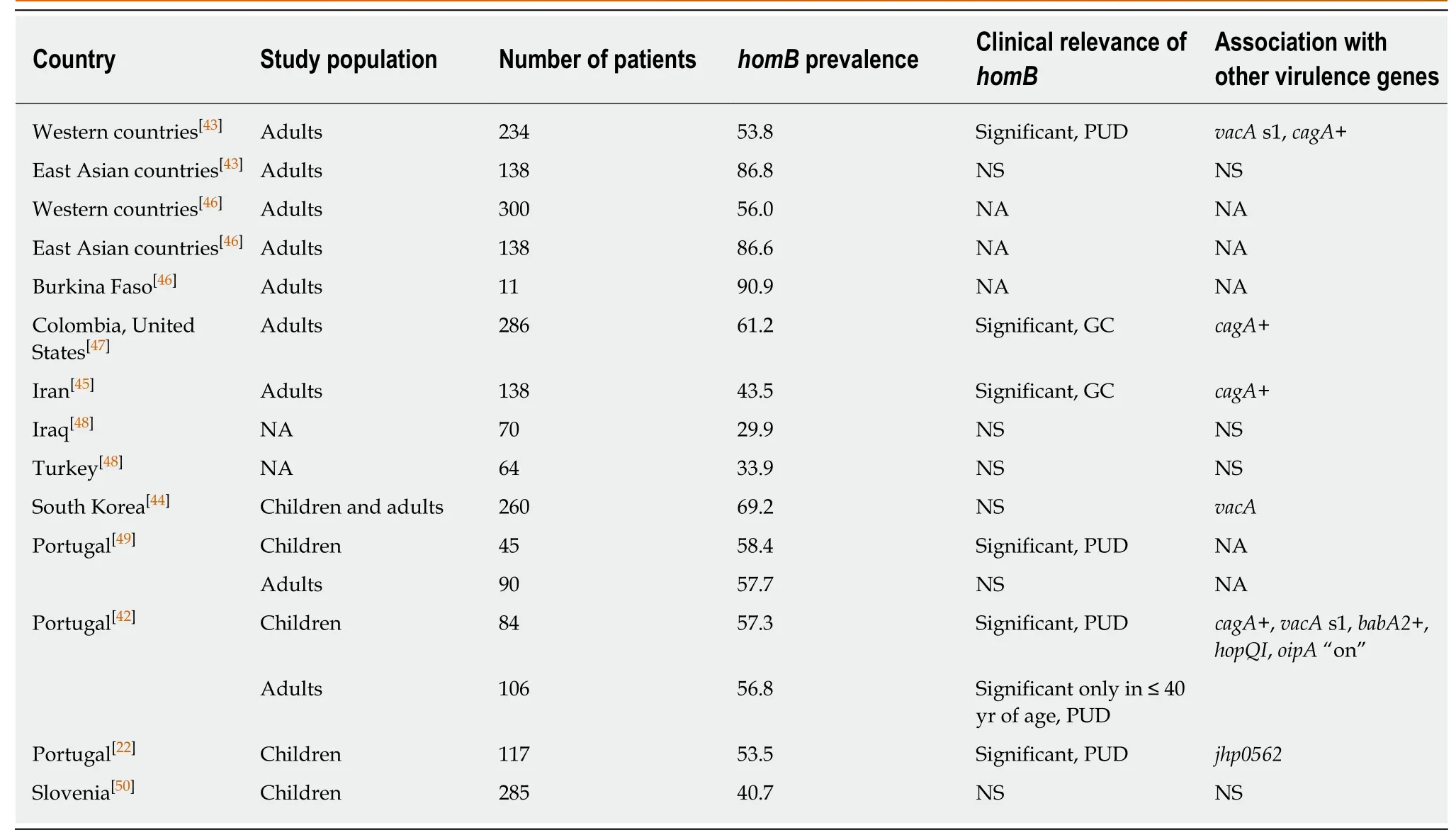
Table 2 Overview of studies on homB prevalence and clinical significance in adults and children
In a study on 200 patients from Colombia and the United States, sabA “on” status was shown to be associated with the presence of pre-neoplastic lesions (e.g., gastric atrophy and severe intestinal metaplasia) and GC. Moreover, sabA “on” was the only predictor of GC versus duodenal ulcer (OR = 2.8, 95%CI: 1.2-6.7) among several investigated OMPs in this study[36]. However, there were no statistically significant differences among Taiwanese patients with sabA “on” and sabA “off” in terms of the prevalence of gastric atrophy or intestinal metaplasia[14]. Although all H. pylori isolates from Iranian patients with GC were found to be sabA “on” (5/5, 100%), the link did not appear to be statistically significant[31]. Similarly, there was no correlation between sabA “on” and clinical outcome among Italian and Japanese patients[35,54], although sabA “on” was associated with atrophy and severe neutrophil infiltration in patients from Japan[54].
Children:In children, the prevalence of the sabA “on” genotype was found to be 44.0% among strains from Portugal and the sabA “on” status significantly correlated with NUD (P = 0.028, OR = 0.298)[42]. Similarly, a low rate of SabA producing strains(38.0%) was detected in a collection of gastric biopsies from children and young adults[55]. Interestingly, it has recently been proposed that high expression of sabA may be responsible for iron deficiency anemia in children and young adults[56].
Associations with other virulence genes:Studies evaluating associations between sabA and other virulence genes are somewhat contradictory. Whereas sabA was closely related to cagA and babA2 positivity in European strains[52], subsequent studies could not confirm these findings[36,37].
Comment:Again, identification of the sabA “on” status by using PCR and sequencing may not reliably reflect the actual production of SabA, thus affecting the result interpretation of studies on sabA clinical relevance, which have primarily used sequencing-based methods[3,14].
VIRULENCE GENES THAT PRODUCE TOXINS AND CAUSE HOST TISSUE DAMAGE
cagA, cagPAI and EPIYA motifs
It has been previously shown that highly virulent H. pylori strains harbor the cytotoxin-associated genes pathogenicity island (cagPAI), which is a 40 kb region containing 31 genes that encode for components of a type IV secretion system,involved in CagA translocation and the host's inflammatory response[4]. cagA is arguably the most extensively studied H. pylori virulence gene to date. It is located at the end of the cagPAI and encodes a 120-145 kDa immunodominant protein, CagA[57].Based on CagA production, H. pylori isolates can be divided into two groups: cagA negative and cagA positive. During infection, CagA is localized on the plasma membrane, where it is phosphorylated at specific Glu-Pro-Ile-Tyr-Ala (EPIYA)-motifs by host Src and Abl kinases. Four distinct segments harboring EPIYA-motifs have been described so far, designated as segments A, B, C, and D[11,57,58]. The biological activity of CagA depends on the number and types of the EPIYA-motifs at the Cterminal region. Following translocation, CagA interacts with multiple host cell molecules and is responsible for dysregulation of homeostatic signal transduction of gastric epithelial cells, induction of pro-inflammatory responses that lead to chronic inflammation of gastric mucosa, and induction of carcinogenesis through the modulation of apoptosis, disruption of cell polarity and promotion of genetic instability. Hence, due to its cancer-inducing traits, CagA was designated as the first bacterial oncoprotein[57,59].
Adults:An analysis of a global collection of H. pylori strains from 53 different geographical/ethnic sources showed the presence of cagPAI in more than 95% of strains from Western and South Africa and East and Central Asia, whereas the presence of cagPAI in other regions ranged from 81% (Northeastern Africa) to only 28% (Latin America). The prevalence of cagPAI in Europe was shown to be intermediate, with approximately 58% of strains harboring cagPAI[60]. The prevalence of cagA positive strains is approximately 60% and > 90% in Western and Asian countries, respectively[2]. In the Middle East, cagA is detected in nearly half of the strains[61].
Since the majority of East Asian strains harbor cagA irrespective of the disease status, it cannot be considered a useful marker of the disease. Nevertheless, based on mosaicism within the EPIYA-motifs, cagA positive strains can be further divided into Western (EPIYA-ABC, EPIYA-ABCC and EPIYA-ABCCC) and East Asian strains(EPIYA-ABD)[5,62]. Although very rarely, a subset of East Asian strains can possess a Western type EPIYA motif, whereas the reverse is not true for Western strains[32,58]. In Latin America, EPIYA-ABC is the most common motif, detected in approximately 51.6%-73.6% of strains, although strains with multiple EPIYA-C segments were found to be rare (2.7%) in a Venezuelan population[32].
When assessing the risk of infection with cagA positive strains for the development of GC, one must be aware of the considerable global variation, not only in the prevalence of cagA positive strains but also in the incidence of GC[60,63]. In Western countries, the presence of cagA is associated with a higher risk of GC and PUD development, whereas in East Asia, where almost all H. pylori strains contain cagA,this association is evident but less prominent[5]. Specifically, patients infected with H.pylori who had CagA antibodies were shown to have a 5.8-fold (95%CI: 2.6-13.0)increase in the likelihood of developing GC compared to uninfected individuals,whereas those who were CagA seronegative only had a slightly but not statistically significantly (OR 2.2, 95%CI: 0.9-5.4) increased risk of GC[64]. Moreover, a metaanalysis of CagA serostatus performed on 10 non-cardia gastric cancer case-control studies from Western populations showed marked differences in CagA seropositivity in H. pylori infected cases (62.8%, n = 1707) and controls (37.5%, n = 2124), with CagA seropositive status associated with a higher risk of GC development (OR = 2.87,95%CI: 1.95-4.22) compared to the risk of being infected with H. pylori only (OR = 2.31,95%CI: 1.58-3.39)[65]. Similarly, a meta-analysis of 10 gastric cancer case-control studies from East Asia also identified an association between CagA seropositivity and increased risk of GC[66], although OR (OR = 1.81, 95%CI: 1.30-2.11) was lower compared to that of Western populations[65,66]. In addition, a large meta-analysis on more than 17000 individuals identified a 1.69-fold risk (95%CI: 1.12-2.55) of PUD among cagA positive Western and Asian populations, with an even higher risk of GC(OR = 2.09, 95%CI: 1.48-2.94)[67]. CagA is also one of the few virulence factors associated with the development of gastric high-grade B cell lymphoma[11].
Different diagnostic approaches should be applied in different geographical regions—due to the almost universal presence of the cagA gene in East Asian strains,the sensitivity of cagA gene detection is suboptimal, rendering cagA subtyping in order to identify those with high risk infections[11]. The number of EPIYA segments in the second repeat region is thought to be associated with GC. Namely, initial trials showed that the incidence of GC was considerably higher if patients were infected with strains harboring multiple EPIYA-C segments (EPIYA-ABCCC) than if patients were infected with strains harboring only one EPIYA-C segment (EPIYA-C).Unfortunately, because East Asian strains only harbor a single EPIYA-D segment,differentiation between chronic gastritis and GC using only the number of repeat regions has proved to be somewhat problematic[5,62]. To clarify this issue, a recent meta-analysis evaluated the differences in PUD and GC risk among strains carrying one EPIYA-D motif or multiple EPIYA-C motifs. In Asian strains, the presence of one EPIYA-D motif was significantly associated with increased GC risk (OR = 1.91, 95%CI:1.19-3.07) compared with the presence of one EPIYA-C motif, whereas it was not significantly associated with PUD (OR = 0.90, 95%CI: 0.46-1.76). Moreover, multiple EPIYA-C motifs were associated with increased PUD risk (OR = 2.33, 95%CI: 1.29-4.20) in Asian countries and with increased GC risk (OR = 3.28, 95%CI: 2.32-4.64) in Western countries[68].
Children:In children, cagA is the best characterized among all virulence genes.Similar to adults, the prevalence of cagA in children varies among different countries/regions. The cagA gene can be found in more than half of H. pylori isolates obtained from symptomatic children from Western countries, namely 60.8% in Poland[18], 59.6% in Slovenia[69]and 70.0% in United States[41]. A surprisingly low prevalence of cagA was found in Portuguese children (22.4%)[21]. In Iran, the reported prevalence of cagA in symptomatic children ranges between 60.0 and 72.7%[70,71]and is similar to that in Turkish children (55.6%-61.0%)[25,72]. A high prevalence of cagA(73.0%) was also observed in symptomatic Venezuelan children with recurrent abdominal pain[73]. In Mexican children, cagA and cagPAI were detected in 63.3% and 71.4% of strains, respectively[74]. Similar to adults, strains from Korean and Japanese children almost exclusively carry the cagA gene (94.0% and 100%, respectively)[75,76].Interestingly, it has previously been shown that the prevalence of cagA can be surprisingly high (66.1% and 75.0% in Colombia and Brazil, respectively) in asymptomatic children from high-risk populations, with rates that are comparable or even higher than those in symptomatic children from other regions[30,77]. It is thus possible that the high prevalence of virulent H. pylori variants in Colombian and Brazilian children contributes to the increased GC incidence in adults from the same region[77]. The high proportion (40.0%) of strains with multiple EPIYA-C motifs further confirms previous observations that this population may already be exposed to the most virulent variants of H. pylori at a young age[30]. The fact that infection with H.pylori is a risk factor for GC highlights the importance of early detection of H. pylori virulence factors in children, especially those residing in areas with a high prevalence of GC[77].
In China, the rates of cagA positivity in the pediatric population closely resemble those in adults, with the prevalence of cagA among children with symptomatic gastroduodenal disease being 94.4%, with no clinical relevance[78]. Similarly, because the cagA positive genotype is present in virtually all Korean and Japanese pediatric strains, no associations with severity of gastritis or PUD were found[75,76]. In contrast,cagA was significantly associated with PUD (OR = 14.06, 95%CI: 4.78-41.29)[42], higher H. pylori density score, and the degree of chronic and acute inflammation[69]in European children.
Associations with other virulence genes:Interestingly, almost all vacA s1 strains also carry cagA, whereas almost all cagA negative strains harbor the less virulent genotype vacA s2/m2[69,79]. In addition, cagA is also more commonly detected in babA2 positive strains[77].
Vacuolating cytotoxin A
The vacuolating cytotoxin A (VacA) derives its name from its capacity to induce the formation of vacuoles in eukaryotic cells. Several other cellular functions of VacA with a potential influence on host cell death have been described thus far, including disruption of endocytic trafficking, release of organic anions and HCO3, promotion of immune tolerance and chronic infection through inhibition of various immune cells,activation of mitogen-activated protein kinases, and modulation of autophagy[80,81]. All H. pylori strains carry the vacA gene, although with different vacuolating ability,which is conferred by variations in five vacA regions: s-region (s1 and s2), i-region (i1,i2, i3), m-region (m1 and m2), d-region (d1 and d2), and the recently identified cregion (c1 and c2). The vacA s2 variant is considered less pathogenic than the s1, since VacA s2 toxins are produced and secreted at lower rates and are also unable to form membrane channels through which VacA s1 induces vacuolation of cells[3,79,81]. VacA i1 is also associated with increased activity compared to VacA i2. Unlike VacA m2, VacA m1 induces a decrease in intracellular levels of glutathione and an increase in oxidative stress, leading to autophagy and apoptosis of host cells[81,82].
Adults:The distribution of vacA alleles is geographically dependent, with s1c being the most prevalent allele in East Asia, while the vacA s1a allele is detected more often in Northern Europe and vacA s1b in Portugal and Spain. In Northern America, vacA s1a and vacA s1b are relatively evenly distributed, whereas virtually all strains from Latin America carry vacA s1b. The vacA s1 allele prevalence ranges from 36.0% in North Africa to 95.0% in East Asia. vacA m1 and m2 are equally distributed, except in Portugal, Spain and Latin America, where vacA m1 is more prevalent (86.2%). The vacA m2b allele is found solely in East Asian strains carrying vacA s1c[83]. Interestingly,mixed vacA s1a/s1b/m2 was found to be the most common genotype in Saudi Arabia[61].
Several studies have intensely focused on potential associations between vacA alleles and risk of PUD and GC. Results were relatively consistent, since most studies identified vacA s1, vacA i1 and vacA m1 alleles as being associated with a higher risk of precancerous lesions and GC[67,84]. Interestingly, vacA i1 and d1 were shown to be significantly associated with non-cardia GC (OR = 37.52, 95%CI: 3.04-462.17 and OR =7.17, 95%CI: 1.43-35.94, respectively), but not with cardia GC. The presence of these alleles may also predict the risk according to the GC type, as vacA i1 was linked to intestinal-type adenocarcinoma (OR = 14.04, 95%CI: 2.15-91.77) and vacA d1 to diffuse-type adenocarcinoma (OR = 7.71, 95%CI: 1.13-52.28)[85]. Furthermore, strains harboring vacA s1 and vacA m1 genotypes were also more commonly detected in patients with severe inflammation and gastric epithelial damage and PUD than in those who were vacA s2/m2 positive. In Western countries and the Middle East, the presence of vacA s1/m1 is associated with an increased risk of PUD, whereas in East Asia, the vacA s1/i1/m1 genotype is not a useful differentiating factor since most strains harbor this genotype[11,34,61]. Moreover, a meta-analysis showed that vacA i1 confers higher risk of GC (OR = 5.12, 95%CI: 2.66-9.85), especially among the Central Asian population (OR = 10.89, 95%CI: 4.11-20.88). Conversely, vacA i1 was not associated with increased risk of PUD (OR = 1.38, 95%CI: 0.87-2.17)[86]. As shown by Van Doorn et al[83], the vacA s1/cagA+ genotype is associated with PUD in all regions of the world.
Children:Genotype vacA s1/m2 is the most common genotype in children from Iran(45.5%) and Turkey (57.1%)[25,70]. In Venezuela, 85.0% of strains obtained from symptomatic children harbored vacA s1/m1[73]. In Slovenia, pediatric H. pylori strains more commonly contain vacA s1 and m2 than vacA s2 and m1, with most strains harboring the vacA s1/m1 genotype[17,50,69]. In asymptomatic Brazilian children, vacA s1(82.5%) and vacA i1 (75.0%) were the most common alleles, whereas m1 and m2 were found to be equally distributed (48.2% each)[30]. Using stool samples, the prevalence of the vacA s1 gene in asymptomatic Colombian children was shown to be very high(91.7%) and similar to that in the adult population (93.2%)[77]. Results from Brazil, a high-risk region for GC, also suggest that asymptomatic children from this area are more often colonized with strains harboring the toxigenic vacA s1 allele[87].
In Iranian children, nodular gastritis was commonly found and was significantly associated with the presence of vacA m1[70]. Similar to cagA, vacA s1 has been strongly associated with PUD risk (OR = 14.13, 95%CI: 4.75-42.04) among Portuguese children[42], whereas there were no significant correlations between vacA status and PUD in Iranian children[71]. Moreover, studies on Korean, Japanese and North American children found no associations between the vacA genotype and clinical outcome or severity of inflammation[75,76,88,89].
Associations with other virulence genes:Compared to vacA s2, strains that harbor vacA s1 more commonly contain cagPAI, babA2, homB and oipA “on”[81]. vacA i1 is strongly associated with vacA s1 and vacA m1 and cagA[30,84].
VIRULENCE GENES WITH OTHER FUNCTIONS
Duodenal ulcer promoting gene
The duodenal ulcer promoting (dupA) gene encompasses jhp0917 and jhp0918, located in the plasticity region of the H. pylori genome. Due to its high homology with the virB4 factor, dupA presumably forms a type IV secretion system together with vir genes, although its exact functions are not yet fully understood. The detection of the dupA gene correlates with increased IL-8 production from gastric epithelial cells, both in vivo and in vitro. Increased IL-8 secretion from the gastric antrum thus leads to the development of predominantly antral gastritis, a well-known characteristic of duodenal ulcer disease[90].
Adults:Worldwide, approximately 48.0% of strains carry dupA[91], with the highest rates in Brazil (89.5%) and South Africa (84.8%)[92,93]and lowest in East Asian countries[91]. A study on 500 isolates from patients with gastritis, PUD and GC originating in Japan, Korea and Colombia showed an overall prevalence of dupA of 26.3%[94]. Surprisingly, the prevalence of the dupA gene was higher in Colombia(36.5%) than in Korea (16.8%), regardless of the clinical outcome[94]. In relation to the prevalence in strains from patients with functional dyspepsia, dupA was detected in 65.0%, 37.8%, 35.7%, 28.9% and 7.1% of strains from Swedish, Australian, Malay,Chinese and Indian patients, respectively[95].
Interestingly, in contrast to other virulence factors, such as cagPAI, vacA, oipA and babA2, which are reportedly associated with an increased risk of both PUD and GC,dupA was the first H. pylori virulence factor to be correlated with a differential susceptibility to PUD and GC, with protection against pre-neoplastic lesions and GC(OR for GC = 0.42, 95%CI: 0.2-0.9, compared with gastritis)[94]. However, some subsequent studies failed to reproduce these results. A meta-analysis on the relationship between the dupA gene and clinical outcomes was therefore performed and it showed that infection with H. pylori strains carrying dupA had a 1.41-fold(95%CI: 1.12-1.76) increased overall risk of duodenal ulcer. A subgroup analysis identified higher ORs in Asian countries (OR 1.57, 95%CI: 1.19-2.06) than in Western countries (OR 1.09, 95%CI: 0.73-1.62), suggesting that dupA can be considered a disease-specific virulence factor, especially in Asian countries. No associations between the presence of dupA and GC or gastric ulcer were found[66]. In addition, the same authors reported that the presence of dupA may also be an independent risk factor (OR = 3.71, 95%CI: 1.07-12.38) for H. pylori eradication failure[90]. Interestingly, a recent study showed protective effects of the dupA gene against severe outcome in infected females (OR = 0.05, 95%CI: 0.01-0.42). Moreover, whereas the sole presence of vacA i1 carried the highest risk for a severe clinical outcome, the simultaneous presence of the dupA gene resulted in a delay of severe disease outcome by almost 20 years[96].
Children:The prevalence of dupA was found to be 37.5% in Mexican children with recurrent abdominal pain[74]. In contrast, all H. pylori strains from Brazilian children were found to be dupA positive, with a significantly higher prevalence than in adults from the same region[92]. However, despite using the same primers for detecting the dupA gene as Gomes et al[92], another study analyzing Brazilian children showed a much lower (37.0%) prevalence of this gene[97]. These discrepancies may be due to the presence of significant geographic differences even within the same country/region,variations in studied populations or rearrangements within the plasticity zone, which is prone to frequent change[92,97].
Associations with other virulence genes:The dupA gene has previously been associated with cagA[74,92,97]and cagPAI[74].
COMBINATIONS OF VIRULENCE GENES
Since some genes are almost exclusively associated with one another (e.g., vacA s1/i1/m1 and cagA), it is impossible to consider each of these virulence genes separately as independent markers for disease outcome. For example, the presence of oipA “on” is tightly linked to the presence of cagPAI and some studies even suggest that cagPAI and OipA act synergistically by regulating the signaling pathways that induce inflammation and actin dynamics[29]. Here, we briefly summarize some of the most intriguing combinations of H. pylori virulence genes.
As expected, the risk of a severe clinical outcome increases if multiple virulence genes are simultaneously detected. It has been shown that strains harboring the vacA s1/m1/cagA+ genotype carry a 4.8-fold (95%CI: 1.71-13.5) increased risk of progression of pre-cancerous lesions in comparison to the strains carrying vacA s2/m2/cagA-, with higher ORs than if each of these virulence genes was evaluated individually[98]. In addition, strains carrying cagA, vacA s1 and babA2 were associated with duodenal ulcer and adenocarcinoma[15], whereas cagA, vacA s1/m1 and babA2 were found to work synergistically in causing intestinal metaplasia[27]. Furthermore, a study from Portugal identified an increased risk of PUD in strains that simultaneously harbored homB, cagA and vacA[43]. Using binary logistic regression, cagA+/homB+ and cagA+/vacAs1 genotypes were found to have the highest discriminatory capacity to distinguish PUD from NUD in children, among the evaluated combinations of virulence factors[42]. Another study on pediatric strains showed that quadruplepositive strains (vacA s1/m1/cagA+/babA2+) had the highest discriminating value for detecting the severity of gastritis compared to other groups evaluated[17]. Interestingly,whereas homB was not associated with a severe finding on gastric histology when considered as an individual marker of the disease, a correlation between the vacA s2/m2/cagA-/babA2-/homB+ genotype and the presence of atrophic changes in Slovenian children was found[50]. Moreover, a study evaluating the prevalence and relevance of various H. pylori virulence factors in the pathogenesis of low-grade gastric MALT lymphoma was unable to identify correlations between any of the putative virulence genes and MALT lymphoma when evaluated individually.However, when using multiple correspondence analysis, patients infected with strains carrying iceA1, sabA “on” and hopZ “off” had 10-fold higher odds (OR = 10.3,95%CI: 1.2-86.0) of developing MALT lymphoma than age-matched patients with gastritis[99].
CONCLUSION
H. pylori isolates show a high degree of geographic variability. It is thus possible that certain H. pylori genotypes are associated with a more severe clinical outcome in some regions, while presenting as virtually harmless variants in other studied populations.The observed discrepancies in several studies on H. pylori virulence genes may be due to various factors: different definitions or diagnoses of gastroduodenal disease,limitations of PCR and sequencing methods for detecting virulence genes (e.g.,inadequate PCR primer design, disregarding frameshift mutations that could have a considerable influence on protein expression and/or function, and poor correlation of the genotypic methods with the actual expression profile of the protein), and inability to detect mixed infections with more than one strain at a time. Moreover, differences between East Asian and Western strains confirm the hypothesis that the degree of gastroduodenal pathology depends on complex relationships between host genetics,environmental factors and the presence, as well as combinations, of various H. pylori virulence genes. Although the importance of the majority of H. pylori virulence genes has not yet been uniformly clarified, knowledge on their role in pathogenesis, as well as disease outcome, has substantially improved in the last two decades. Careful monitoring and continuous refining of their roles will not only contribute to novel strategies for H. pylori vaccine development but also impact potential alternative therapies and facilitate the discovery of novel virulence genes. Although sequencing methods have dramatically improved over the years, enabling better and in-depth information on H. pylori genome structure, future studies should not only focus on these methods but also account for differences in protein expression profiles.Nevertheless, enriched knowledge on the pathogenicity of H. pylori virulence genes may be of clinical significance, since the detection of more virulent variants of strains,such as those with an increased number of CagA EPIYA-motifs, could be used to improve clinical prediction of the disease risk and identify those who need more intensive surveillance and eradication of the infection to prevent serious healthrelated consequences. In addition, focusing on a single virulence factor is probably too restrictive, since clear linkages between various virulence factors with different biological roles and significances exist, which may act synergistically to induce serious gastroduodenal pathology. Moreover, in the light of recent studies demonstrating that early exposure to H. pylori provides some protection against subsequent atopy and allergic conditions in childhood[100], identification of reliable discriminative virulence factors of bacterial strains could be extremely helpful in the event that triaging of H. pylori infection is applied in the future.
杂志排行

World Journal of Gastroenterology的其它文章
- Role of NLRP3 inflammasome in inflammatory bowel diseases
- Gastroesophageal reflux disease, obesity and laparoscopic sleeve gastrectomy: The burning questions
- Intestinal permeability in the pathogenesis of liver damage: From non-alcoholic fatty liver disease to liver transplantation
- Crosstalk network among multiple inflammatory mediators in liver fibrosis
- Neoadjuvant radiotherapy for rectal cancer management
- Occupational exposure to vinyl chloride and liver diseases