番茄的配合力和杂种优势群的划分
2019-09-10金兰赵丽萍王银磊徐丽萍周蓉宋刘霞崔霞李仁赵统敏余文贵
金 兰 赵丽萍 王银磊 徐丽萍 周蓉 宋刘霞 崔霞 李仁赵统敏 余文贵




摘要: 本研究筛选出14个番茄亲本,采用Criffing完全双列杂交设计组配105个杂交组合,对13个性状进行统计,分析配合力及杂种优势,划分优势群确定杂种优势。结果表明,F1的杂种优势在大部分的性状中均存在,F1除果实硬度和纵径外的11个性状值均显著大于亲本。亲本11和亲本8的单株产量、单果质量等性状的一般配合力最大,亲本lx亲本11、亲本4x亲本11和亲本1×亲本4的产量性状均具有较高的一般配合力和特殊配合力。配合力与杂种优势相关性分析结果表明,大部分性状杂种优势与特殊配合力显著相关,单株产量的一般配合力与杂种优势显著相关。一般配合力聚类分析和F,的性状表现聚类分析2种方法划分优势类群,结果证明一般配合力聚类分析方法是有效的,它将14个亲本划分为5个优势群,并确定了优势群1( HGI)×优势群2(HC2),优势群1( HGl)x优势群5(HC5)和优势群2(HC2)×优势群5(HC5)这3个杂优模式。本研究揭示了一般配合力分析法可以作为番茄杂交育种优势群划分的一种重要方法,并为番茄杂交育种中亲本的选择提供指导。
关键词:番茄;配合力;杂种优势;杂种优势群;杂优模式
中图分类号:S641.2
文献标识码:A
文章编号: 1000-4440(2019)03-0667-09
番茄是茄科番茄属的草本植物,含有丰富的营养物质(如维生素A、维生素C和番茄红素等),种植面积广,但生产水平仍然不能满足市场需求。提高产量和品质是番茄育种的主要育种目标,而杂交育种对番茄产量和品质的提高具有重要意义。亲本的选择和组配是杂交育种的重要组成部分,分析亲本的配合力并根据配合力来选择亲本和评价杂交组合,是常用的亲本选配方法。研究结果表明,亲本配合力的高低对杂种优势强弱有影响,高产强优势组合的选育不仅跟亲本的一般配合力水平有关,而且与两亲本之间的特殊配合力有关[1-3]。杂种优势利用的理论基础是杂种优势群和杂种优势模式,杂种优势群的划分和杂种优势模式的确定被很多研究结果证明对玉米、水稻等作物的育种具有明显提高育种效率,加速育种进程等作用[4-5],也是杂种优势利用的一种途径。在作物中,杂种优势群的划分在育种中最早得到研究和验证的作物是玉米,美国是最早研究玉米杂种优势群的国家,也是最早形成杂种优势群和杂种优势模式的国家,而有关番茄杂种优势群的划分和杂种优势群的确定的研究很少。
划分优势群的方法有很多,如系谱分析法、数量遗传学方法、生化标记分析法、分子标记法以及多种方法综合利用。国内育种家通常分析种质农艺性状的配合力,根据配合力进行聚类分析,并结合系谱来源、杂种优势来划分杂种优势群和确定杂种优势模式[6-7]。配合力分析法是从遗传学角度对亲本进行分析,将亲本进行双列杂交,选取相应杂种一代的相关性状,计算其一般配合力和特殊配合力,然后根据配合力值进行聚类,划分杂种优势群。Wang等[8]基于分子标记、产量优势和配合力分析将17个水稻亲本材料划分为4个优势群并确定为3个杂种优势模式。虽然每种分析方法都具有局限性,但都为之后杂种优势群的划分提供了很好的参考。
本研究基于前期的324份栽培番茄的遗传多样性分析,筛选14个番茄代表,通过Griffing完全双列杂交[p(p+1)/2]组配,并计算配合力和杂种优势,评价14个栽培番茄的杂种优势和配合力,确定番茄的杂种优势群和杂优模式,为以后番茄的杂种优势群划分提供指导。
1 材料和方法
1.1 材料
本试验选用14个性状不同的栽培番茄材料:228、CLN2498D、AVT01174、9V5F224、H24、BJF3、LH18-330、SY-Cheng、B072、LH18-250、LH18-294、LA2010、31号大红、VFN作为亲本,亲本性状见表1。2017年秋季在江苏省农业科学院六合试验基地播种亲本材料,花期进行人工去雄授粉,按照Griffing完全双列杂交[p(p+1)/2],配制105个杂交组合。
1.2 方法
2018年1月5日,将105个杂交组合和14个亲本材料播种于江苏省农业科学院六合试验基地,于2018年3月9号将番茄苗定植于大棚中,设计为随机区组,2次重复,每个重复10株,每个小区2行,常规管理。在番茄生长期,依据《番茄种质资源描述规范和数据标准》[9]统计熟性性状(始花期、坐果期和始收期),产量性状(单果质量、单株产量、第一穗坐果数、纵横径)和果实品质(果实硬度、果肉厚度、可溶性固形物和酸含量)。
1.3 数据处理
数据处理采用SPSS v16.0和Microsoft OfficeExcel软件。
2 结果与分析
2.1 亲本和Fi的性狀
14个亲本与105个杂种一代的主要农艺性状表现如表2所示。13个性状中除了硬度和纵径,F1的表现值都大于亲本。其中,F1的单株产量,单果质量,果肉厚度和可溶性固形物含量明显大于亲本;F1的熟性值(始花期,坐果期和始收期)和首花节位与亲本差异不明显。结果表明番茄亲本杂交能有效改良单株产量,单果质量,果肉厚度和可溶性固形物含量,其中硬度的降低可能与可溶性固形物含量的提高有关。
2.2 F1主要性状杂种优势
从表3可以看出,番茄的13个性状的超亲优势普遍存在,无优势的占比很少。除始收期、硬度和纵径外的10个农艺性状的正向超中优势比例大于负向超中优势;番茄的单株产量和第一穗坐果数的正向超高优势的占比大于负向超高优势。不同性状的杂种优势程度不同,对各性状的平均超中优势值进行排序,得到:单果质量>单株产量>第一穗坐果数>果肉厚度>横径>首花节位>酸含量>可溶性固形物含量>始花期>坐果期>始收期>纵径>硬度;超高优势的强弱顺序为:单株产量>第一穗坐果数>酸含量>单果质量>始花期>果肉厚度>可溶性固形物含量>横径>首花节位>始收期>坐果期>硬度>纵径。单株产量超中优势值和超高优势值靠前,说明单株产量的杂种优势明显,并且获得正向杂种优势比较容易;纵径和硬度的超中优势和超亲优势都靠后,说明纵径和硬度的负向杂种优势明显。番茄F1的13个性状的超中优势中,10个性状为正向超中优势,占总体性状的76.92%,2个性状为负向超高亲优势,占比15.38%,这表明10个性状要获得具有正向超中优势的F1比较容易,而获得正向超高亲优势需要特殊亲本的选配。从标准差可以看出,13个性状杂种优势的变异幅度较大,其中第一穗坐果数超中优势和超高优势标准差最大,表明杂交组合第一穗坐果数杂种优势上表现出最大差异:坐果期和始收期的超中优势和超高优势标准差靠后,表明杂交组合在坐果期和始收期的杂种优势上变异小。
2.3 番茄主要性状的方差分析
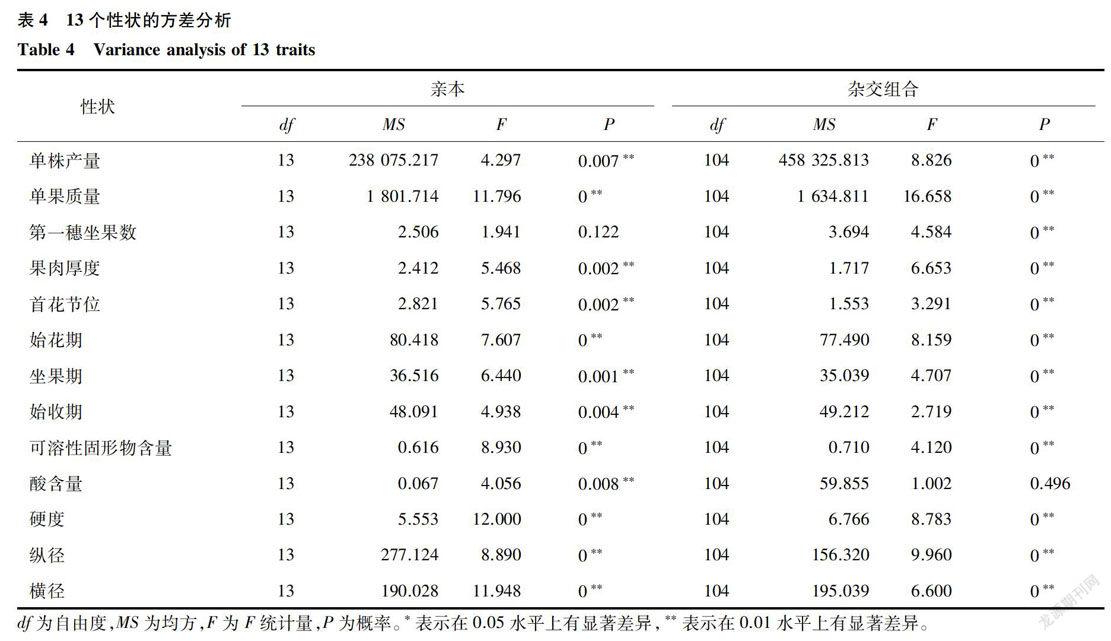
从表4可知,亲本性状中,只有第一穗坐果数F值没有显著性,其他性状都达到了显著水平,说明调查的13个性状中除了第一穗坐果数外均在亲本间存在差异。从杂交组合来看,只有酸含量的F值没有显著性,说明除了酸含量以外其他12个性状中杂交组合间存在着真实的遗传差异,可以按照Griffing2固定模型和随机模型进行配合力分析。
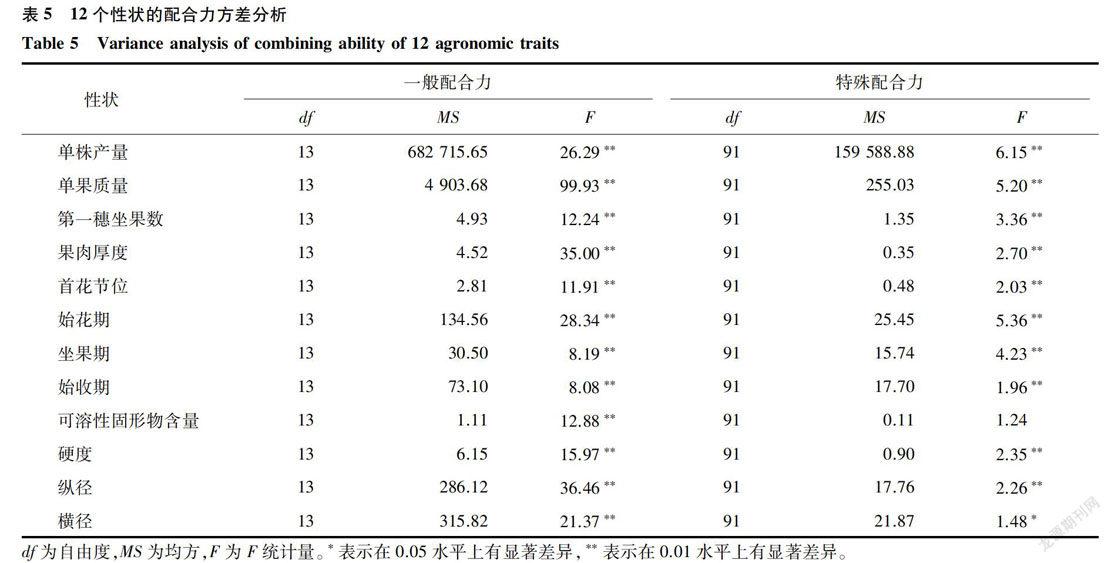
从表5可以看出,杂交组合间的可溶性固形物含量的特殊配合力没有显著性差异,其他性状的一般配合力和特殊配合力均为显著或极显著差异,说明除了可溶性固形物含量的特殊配合力外,其他性状的特殊配合力都存在真实差异,需要进一步做配合力效应分析。
2.4 性状的一般配合力分析和特殊配合力分析
从表6可以看出,亲本11和亲本8的单株产量、单果质量、始花期、坐果期、始收期和横径的一般配合力值均最大,说明亲本11用于番茄提高产量,延迟生殖生长和增大横径的效果最好;亲本13的果肉厚度和纵径的一般配合力大,说明亲本13用来提高番茄果肉和纵径的效果最好;亲本3的硬度和可溶性固形物含量的一般配合力值最大,说明亲本3对提高番茄的硬度和可溶性固形物含量的效果最好。亲本1的坐果期和始收期的一般配合力为最小,说明亲本1对选配早熟品种效果最好。同时可以看出,不同性状,亲本一般配合力效应的差异显著性不同,其中亲本l一般配合力效应的差异显著性最好,亲本9-般配合力效应的差异显著性最差。
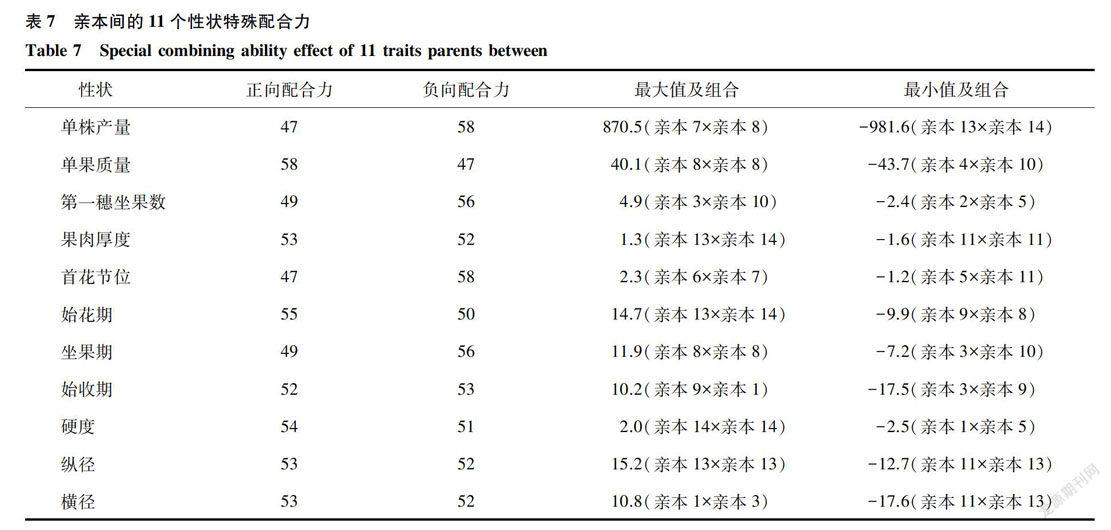
从表7可以看出,单株产量的特殊配合力中,组合亲本7x亲本8最好,并且亲本8的特殊配合力较高,所以此组合为较好的番茄材料。在选育番茄品种时,以选择一般配合力较高的前提下选择高特殊配合力为准则,提高番茄单株产量的较好组合还有:亲本lx亲本11,亲本4x亲本11,亲本1×亲本4,亲本1×亲本13,亲本1×亲本14,亲本4x亲本8,亲本4x亲本13,亲本4x亲本14。单果质量的一般配合力和特殊配合力均较高的组合为:亲本1×亲本4,亲本1×亲本11,亲本4x亲本7,亲本4x亲本11,亲本6x亲本7,亲本6x亲本8和亲本11×亲本13。果肉厚度的配合力中,亲本13和亲本14最大,并且亲本13的一般配合力最大,亲本14其次,所以亲本13和亲本14是提高番茄果肉厚度最好的选育亲本。
2.5 配合力与杂种优势的关系
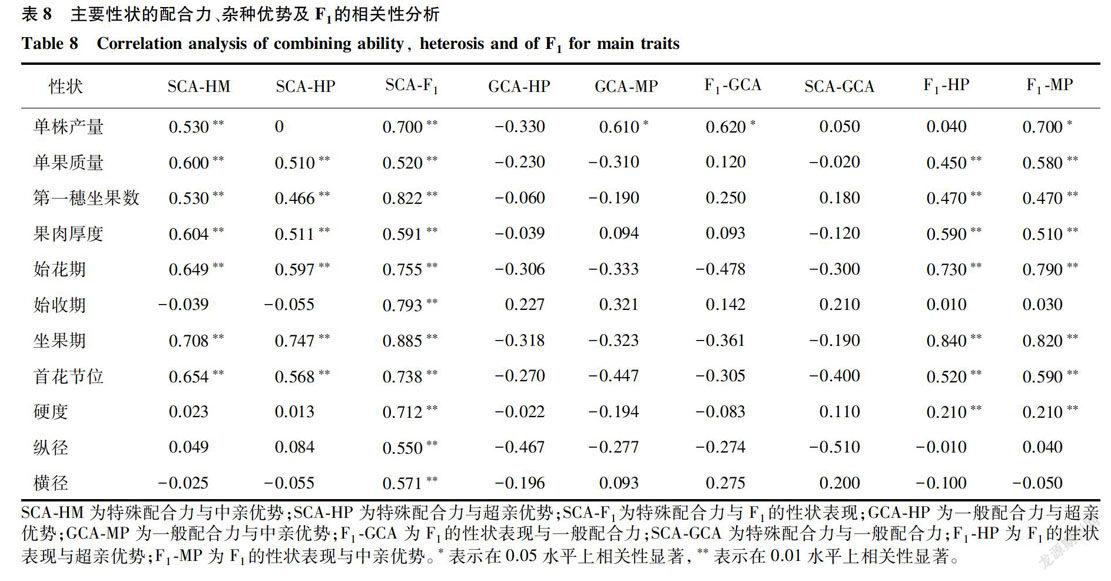
一般配合力、特殊配合力与中亲优势、超高亲优势及F1的表现之间的相关性从表8可知。单株产量的一般配合力和超中优势的相关性很高;大部分性状杂种优势与特殊配合力相关性很高,与组合亲本8x亲本10和亲本2x亲本3的单果质量的超高优势和超中亲优势均较高的结果相符,并且具有较高的特殊配合力;11个性状的一般配合力和特殊配合力间的相关性均不显著,如亲本13和亲本8的单株产量和单果质量的一般配合力均较大,而他们的特殊配合力却为负值;大部分性状的杂种优势与F1的表现显著相关。
2.6 杂种优势群划分
根据主要性状配合力、杂种优势及F,的性状表现相关性分析,F1与特殊配合力及杂种优势显著相关,与一般配合力相关不显著,我们选择亲本的一般配合力和F1的表现值进行杂种优势群的划分。
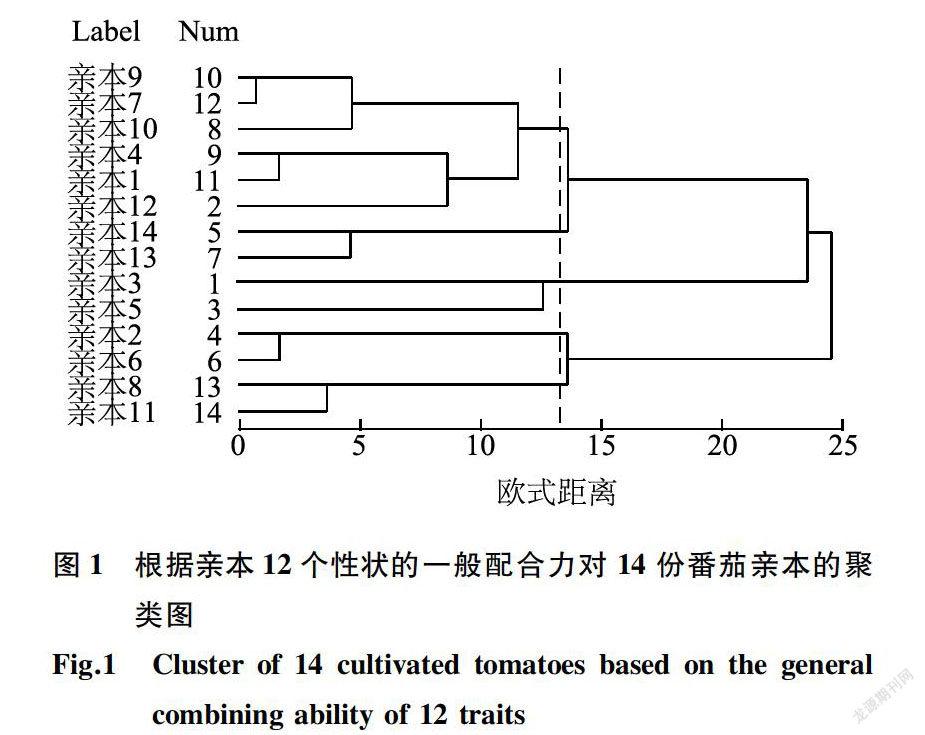
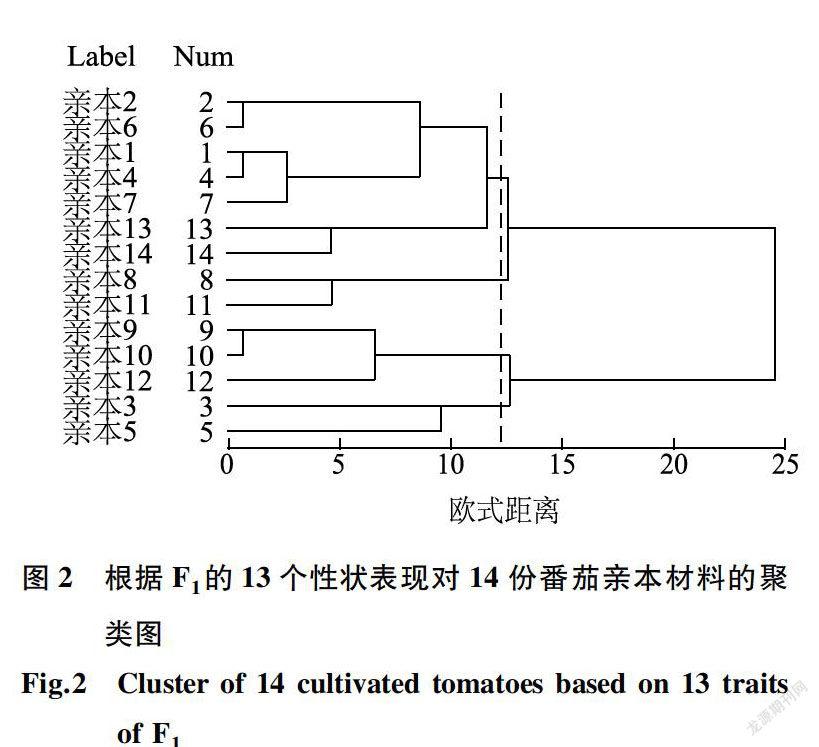
根据14个亲本单株产量、单果质量、第一穗坐果数、果肉厚度等12个性状的一般配合力效应值用SPSS进行标准化后计算欧式距离,采用Ward方法对14个亲本进行聚类分析(图1)。根据14个番茄親本组配的F1表现的单株产量、单果质量、第一穗坐果数等13个性状的平均表现值进行标准化后计算欧式距离,采用Ward方法对14份亲本进行聚类分析(图2)。以13为值域时,一般配合力聚类分析法将14个番茄亲本划分为5大类群,分别为HGI(亲本9、亲本7、亲本10、亲本4、亲本1和亲本12),HG2(亲本14和亲本13),HG3(亲本3和亲本5),HG4(亲本2和亲本6)和HG5(亲本8和亲本11);当值域为12时,Fi的平均表现聚类将14个番茄亲本划分为4大类群,分别为SGI(亲本2、亲本6、亲本1、亲本4、亲本7、亲本13和亲本14),SG2(亲本8和亲本11),SG3(亲本9、亲本10和亲本12)和SG4(亲本3和亲本5)。
3 讨论
有关番茄的杂种优势利用和配合力效应的研究很多[10-12]。杂种优势在作物中广泛存在,并且杂种优势利用作为提高作物产量和品质的一条重要途径,在很多作物上得到广泛的研究和应用。确定杂种优势群是杂种优势利用的关键13],因此本研究通过来自不同类群的14个自交系间杂交,比较杂种优势和配合力,进而划分优势群和确定杂种优势模式。
本研究通过对105个组合和14个亲本的13个性状进行对比分析,结果表明,杂种F1的不同性状均不同程度地表现出优于亲本,与105个杂交组合杂种优势分析结果相符。其中单株产量的杂种优势最好,最大超中优势值和超高优势值分别达到2.51(亲本1×亲本13)和1.71(亲本7x亲本14),高于前人研究结果[14-15]。Aisya等[12]在44个番茄组合的8个农艺性状的杂种优势研究中发现组合的单株产量的最大超中优势和超高优势分别为58.8%和42. 2%。自花植物进行杂交育种的标准是杂种优势值大于等于20%。本研究满足标准的性状有:单株产量、单果质量、第一穗坐果数、酸含量和硬度,与Hannan等[14]和Aisya等[12]的研究结果相符。杂交组合的始花期、坐果期和始收期均表现出弱的杂种优势,其中他们的超高亲优势均小于0,说明亲本杂交对于促进早熟有效果,此结果与Ahmed等16]和Singh等[17]提出的番茄在杂交组配中始花期表现出负向超亲优势的研究结果一致[16-17]。本研究中杂交组合的果肉厚度、单果质量、果实纵横径等性状最大超亲优势值均大于Aisya等[12]和Hannan等[14]的研究结果。
不同基因型间的农艺性状存在显著差异是进行该性状配合力分析的前提[18]。本研究中酸含量基因型间没有显著差异,进而对其他12个性状进行配合力方差分析。大部分性状的配合力都具有显著差异,表明加性效应和非加性效应都对调控除可溶性固形物含量外的11个农艺性状有着重要作用。而Kumar等[19]评估番茄一般配合力和特殊配合力的研究结果显示,单株产量、开花期、单果质量、可溶性固形物和酸含量的配合力具有显著性差异;Aisya等[12]对番茄的产量和产量相关性状的配合力显著性分析结果表明除果肉厚度外其他所有性状配合力都受加性效应和非加性效应调控。
分析14个亲本材料的配合力效应是本研究的重要目的。亲本11的单株产量和单果质量的一般配合力效应为最大,分别为337. 51和36. 01,均高于Akram等[20]的研究结果,说明亲本11杂交比较容易得到较高产量的后代组合。Aisya等[12]的研究中显示其亲本材料的单株产量、单果质量和果实纵横径的一般配合力最大值分别为482. 39、10. 35、5.85和4.25,均小于本研究结果。
配合力与杂种优势的关系比较复杂,夏美翠、王明理等[21-22]研究认为配合力与杂种优势密切相关。本研究结果表明,单株产量的超中优势与GCA显著正相关,Yustiana[23]提出GCA和杂种优势均受主效基因影响,它们之间呈正相关;大部分的农艺性状特殊配合力均与超亲优势显著相关。有关SCA与GCA的关系以往的研究结论各不相同,王明理等[22]认为,双亲一般配合力与特殊配合力没有必然联系,2个高GCA的亲本所配的杂交组合特殊配合力不一定高,与本试验的研究结果相符。
杂种优势群的划分和杂优模式的建立是本研究的另一重要目的,对提高番茄育种效率、加速育种进程有着很大的意义。有关番茄杂种优势群划分和杂优模式确立的研究很少,但水稻、小麦等作物关于这方面的研究比较成熟。陈晓文等[1]利用普通小麦亲本8个性状的一般配合力将15份亲本划分为5类,刘旺清[24]根据不同农艺性状的一般配合力大小将15份普通小麦的亲本划为两大群四亚群。本研究根据一般配合力和F1的性状表现采用聚类分析法划分优势群,分别划分为5个和4个类群,不同的类群番茄种质的表型具有一定的差异。由于有效的优势群划分方法可以使来自不同优势群的2个亲本组配的杂交组合优于来自同一优势群内两亲本组配的组合[25],因此我们对2种划分方法得到的类群群内和群间的杂交组合的性状表现进行对比。一般配合力聚类分析得到的5个类群群间的杂交组合的单株产量、单果质量、第一穗坐果数等9个性状平均表现都显著高于来自群内的杂交组合;群间杂交组合的始花期、坐果期和始收期这3个性状表现优于群内组合,即不同群间杂交可以获得早熟杂交组合;只有群内和群间组合的可溶性固形物含量和酸含量平均表现差异不大,这也与组合间的可溶性固形物和酸含量方差不显著这一结果相符。F1的性状表现聚类分析法得到的4个类群,只有可溶性固形物含量、酸含量、始花期和单株产量这4个性状的群间组合平均表现优于群内组合表现,所以一般配合力分析法优于F1的性状表现聚类分析法。因此一般配合力聚类分析法为有效的优势群划分方法,并将14个栽培番茄材料划分为5个优势群。确定杂种优势模式是本研究的另一重要目的,Lu等'26]研究认为杂优模式可以获得具有较强杂种优势的2个优势群间的组配方式。除始收期、坐果期和始花期外的11个性状的超中优势平均值大于15%的组合按大小顺序排列有:HGlxHG2,HG2 xHG5,HG2 xHG4,HG2×HG3和HGlxHG5;其超高优势大于0的组配方式有:HGlxHG2,HGlxHG5和HG2xHG5,其中HG1×HG2产量的杂种优势最好,HG2 xHG5为其次。所有组配方式的始收期、坐果期和始花期都为负向杂种优势,说明5个優势群间杂交都容易获得较亲本早熟的组合。HG1×HG2、HG1×HG5和HG2×HG5确定为3个杂交优势模式,对番茄杂交育种尤其是提高产量具有很大帮助。
参考文献:
[1] 陈晓文,马守才,王志军,等.15个化杀杂交小麦亲本配合力和杂种优势群的初步研究[J].麦类作物学报,2011,3l(4):630-636.
[2]蒙成,吴雅芳.早熟玉米白交系主要农艺性状配合力分析[J].南方农业学报,2018,49(3):424-430.
[3]宋希云.黄铁城,张爱民,等,杂种小麦强优势组合选配规律的研究Ⅱ,配合力分析[J].北京农业大学学报,1993,19(增刊):45-50.
[4]XIE F M, HE Z Z,ESCUERRA M Q,et al.Determination of het-erotic groups for tropical Indica hybrid rice germplasm[J].TheorAppl Cenet, 2014, 127:407-417.
[5]SUWARNO W B,PIXLEY K V.PALACIOS R,et al.Formationof heterotic groups and understanding genetic effects in aprovitamin A biofortified maize breeding program[J]Crop Sci,2014. 54:14-24.
[6]祁志云,杨华,邱正高,等.几个主要玉米白交系的配合力及聚类分析[J].西南大学学报,2010,32(2):19-25.
[7]李明顺,张世煌,李新海,等,根据产量特殊配合力分析玉米自交系杂种优势群[J].中国农业科学,2002,35(6):600-605.
[8]WANC K,QIU F L,WENCESLAO L.et al-Heterotic groups oftropical indica rice germplasm[J].Theor Appl Cenet, 2015, 128:421-430。
[9] 李锡香,杜永臣,冯兰香,等.番茄种质资源描述规范和数据标准[M].北京:中国农业出版社,2006.
[10]王先裕,于分弟,梁聪耀,等,番茄核雄性不育系大果实硬度杂种优势及遗传效应的研究[J].中国蔬菜,2010( 20):27-31.
[11]田园,张喜春,谷建田,等.番茄亲本完全双列杂交后主要品质性状配合力及遗传力分析[J].中国农学通报,2015,31( 19):87-91.
[12] AISYA S I,WAHYUNI S,SYUKUR M.et al-The estimation ofcombining abilitv and heterosis effect for vield and yieldcomponents in tomato( Solanum lycopersicum MiIL) at Lowland[J] .Journal of Crop Breeding and Cenetics, 2016,2(1):23-29.
[13] MELCHINCER A E, GUMBER R K, JACK E S,et al.Overviewof heterosis and heterotic groups in agronomic crops[J].CSSA,1998, 125:29-44.
[14]HANNAN M M,BISWAS M M.Combining ability analysis of yieldand yield components in tomato(Solanumlycopersicum Mijl)[J].Turk J Bot,2007, 31:559-563.
[15] AHMAD S,OUAMRUZZAMAN A K M. Estimate of heterosis intomato(Solanmlycopersicum L)[J].Bangladesh J Agril Res,2011, 36(3):521-527.
[16] AHMED S U. SHAHA H K, SHARFUDDINA U, et al-Study ofheterosis and correlation in tomato[J].Thai Journal of AgriculturalScience, 1988,21(2):117-123.
[17] SINGH R K, SINCH V K.Heterosis breeding in tomato(Lycoper-sicon esculenturm MilL)[J].Annals of Agril Res, 1993, 14(4):416-420.
[18] SINGH R K. CHAUDHARY B D.Biometrical methods in quanti-tative genetic analysis[M].New Delhj: Kalyani: 1979.
[19]KUMAR R,SRIVASTAVA K.Combining ability analysis for yieldand quality traits in tomato(Solanum lycopersicum L.)[ J].Journal of Agricultural Science, 2013,5(2):213-218.
[20] AKRAM F,HOSSEIN N.The estimate of combining ability andheterosis for yield and yield components in tomato(Lycopersicon es-culentum Mijl.)[J].J Biol Environ Sci, 2012,6(17): 129-134.
[21]夏美翠,宋希云,李储学,等.普通小麦亲本配合力与杂种优势关系研究[J].莱阳农学院学报,1997,14(1):11-15.
[22]王明理,张爱民,黄铁城,等.T型杂交小麦品质及农艺性状的研究I.杂种优势和配合力[J].作物学报,1985. 11(3): 146-158.
[23]YUSTIANA. Combining ability and heterotic group analysis of sev-eral tropical maize inbred lines from PT[ D]. Bogor: Bogo Agricul-tural University, 2013.
[24]刘旺清,小麦轮回选择后代与普通小麦品种间杂种优势的研究[D].北京:中国农业大学,2005.
[25] FAN X M. ZHANG Y M. YAO W H, et al-Classifying maize in-bred lines into heterotic groups using a factorial mating design[J].Agron J, 2009, 101: 106-112.
[26] LU Z M, XU B Q.On significance of heterotic group theory in hy-brid rice breeding[J].Rice Science, 2010. 17(2):94-98.
(責任编辑:陈海霞)
