基于网络药理学探讨丹参酮IIA治疗脊髓损伤的可能机制及相关通路
2019-09-10贺丰朱立国赵赫姚生莲曹峥杨永栋李学朋
贺丰 朱立国 赵赫 姚生莲 曹峥 杨永栋 李学朋

摘要 目的:基于網络药理学分析丹参酮IIA治疗脊髓损伤的分子生物学机制及相关通路。方法:利用TCMSP、BATMAN-TCM、DTPS、STITCHI、Swiss Target Prediction 5个数据库筛选出丹参酮IIA的作用靶点,利用OMIM、GeneCards、CTD数据库检索脊髓损伤相关基因,利用韦恩图筛选丹参酮IIA治疗脊髓损伤的作用靶点。利用STRING数据库进行蛋白互作网络分析,利用Cytoscape软件构建蛋白相互作用网络,利用Metascape数据库进行GO分析和KEGG信号通路富集分析。结果:丹参酮IIA共有潜在靶点182个,与脊髓损伤相关的靶点157个,其中肿瘤蛋白p53(TP53)、Myc原癌基因蛋白(MYC)、G1/S-特异性周期蛋白-D1(CCND1)、半胱氨酸天冬氨酸蛋白酶3(CASP3)、JUN原癌基因蛋白(JUN)5个靶点在蛋白互作网络中具有较为重要的作用。丹参酮IIA治疗脊髓损伤涉及的生物学过程主要包括药物反应、脂多糖反应、细胞对DNA损伤刺激的反应等;涉及的分子功能主要包括酶结合、转录因子结合、蛋白结合等。其作用机制可能与cAMP signaling pathway、MAPK signaling pathway、PI3K-Akt signaling pathway、HIF-1 signaling pathway、TNF signaling pathway、Apoptosis等通路有关。结论:丹参酮IIA治疗脊髓损伤具有多靶点、多通路作用的特点,其可能通过促进神经元存活、促进轴突再生、抑制神经细胞凋亡、抗炎等过程对脊髓损伤过程进行干预。
关键词 丹参酮IIA;脊髓损伤;网络药理学;生物信息学
Exploring the Possible Mechanism and Related Pathways of Tanshinone IIA in the Treatment of Spinal Cord Injury Based on Network Pharmacology
He Feng1,Zhu Liguo1,Zhao He2,Yao Shenglian3,Cao Zheng2,Yang Yongdong4,Li Xuepeng1
Abstract Objective:To analyze the molecular biological mechanism and related pathways of spinal cord injury by tanshinone IIA based on network pharmacology.Methods:The target sites of tanshinone IIA were screened by TCMSP,BATMAN-TCM,DTPS,STITCHI and Swiss Target Prediction.The genes related to spinal cord injury were searched by OMIM,GeneCards and CTD databases,and the target of the spinal cord injury was screened by Wayne map.The STRING database was used for protein interaction network analysis.The protein interaction network was constructed by Cytoscape software,and the Metascape database was used for GO analysis and KEGG signal pathway enrichment analysis.Results:Tanshinone IIA had a total of 182 potential targets and 157 targets related to spinal cord injury,including tumor protein p53(TP53),Myc proto-oncogene protein(MYC),and G1/S-specific cyclin-D1(CCND1).The 5 targets of caspase 3(CASP3)and JUN proto-oncogene protein(JUN)played an important role in the protein interaction network.The biological processes involved in the treatment of spinal cord injury by tanshinone IIA mainly include drug reaction,lipopolysaccharide reaction,and cell response to DNA damage stimulation; the molecular functions involved include enzyme binding,transcription factor binding,and protein binding.Its mechanism may be related to cAMP signaling pathway,MAPK signaling pathway,PI3K-Akt signaling pathway,HIF-1 signaling pathway,TNF signaling pathway,apoptosis and other pathways.Conclusion:Tanshinone IIA has multiple target and multi-channel effects in the treatment of spinal cord injury.It may interfere with the process of spinal cord injury by promoting neuronal survival,promoting axon regeneration,inhibiting neuronal apoptosis and anti-inflammatory.
Key Words Tanshinone IIA; Spinal cord injury; Network pharmacology; Bioinformatics; Mechanism; Pathway
中图分类号:R285文献标识码:Adoi:10.3969/j.issn.1673-7202.2019.11.009
脊髓损伤(Spinal Cord Injury,SCI)是由创伤事件引起的,可导致神经功能受损,部分或全部感觉、运动功能丧失,瘫痪、失去肠道和膀胱控制以及性功能障碍的疾病。它不仅对SCI个体,而且对整个家庭都有深刻的心理、社会和经济影响[1]。据最新研究表明2016年全球脊髓损伤的患病率约为368/105,在我国脊髓损伤的患病率约为236/105,约380万人口正在遭受脊髓损伤带来的困扰[2]。脊髓损伤分为原发性和继发性2个阶段,原发性损伤与神经和血管结构的破坏有关,继发性损伤发生在原发性损伤后几分钟至几天内,包括一些列级联式的病理生理变化,包括血管功能障碍、炎性反应、氧化应激、谷氨酸兴奋性毒性、细胞凋亡等[3]。由于原发损伤是不可逆的,因此如何抑制脊髓损伤后继发的病理生理变化,减轻继发性损伤,以实现神经保护和神经再生功能是当今研究的重点。
丹参酮IIA是中药丹参的主要活性成分之一,聚有抗炎、抗凋亡[4]、抗氧化[5]、神经保护[6]、改善微循环[7-8]等作用,但其分子机制研究尚不深入。网络药理学是基于药物通过“多靶点、多途径、多通路”发挥作用的理念而发展起来的新方法[9],它可整合各种药物、蛋白、基因等数据库,采用生物信息学技术分析和构建药物-靶点-疾病的网络相互作用模型,探索药物发挥作用机制,其整体性、系统性的特点非常适合应用于中药单体及复方的研究。本文采用网络药理学方法对丹参酮IIA治疗脊髓损伤的机制进行探讨,并对未来的临床应用和科学研究提供方法依据和理论基础。
1 材料与方法
1.1 丹参酮IIA靶点的预测 通过TCMSP(http://lsp.nwu.edu.cn/tcmsp.php)、BATMAN-TCM(http://bionet.ncpsb.org/batman-tcm/)、DTPS(http://demo.dtps.iprexmed.com/#/index)、STITCHI(http://stitch.embl.de/)、Swiss Target Prediction(http://swisstargetprediction.ch/)五種数据库及文献挖掘方法筛选丹参酮IIA的相关基因靶点。
1.2 SCI相关基因靶点的筛选 通过GeneCards数据库(http://www.genecards.org)、OMIM数据库(https://omim.org/)和DisGeNET数据库(http://www.disgenet.org)检索与SCI相关基因,汇总后删除重复基因。
1.3 丹参酮IIA治疗SCI相关靶点的收集 利用韦恩图将丹参酮IIA靶点基因与SCI相关基因进行比对,交集的部分很有可能是丹参酮IIA治疗SCI的靶点基因。
1.4 蛋白相互作用网络构建与分析 String数据库(https://string-db.org,Version 11.0)[10]是一个收集了大量蛋白相互作用关系的数据库,其中包含了经过实验验证和通过生物信息学方法预测得到的数据,共涉及9 643 763个蛋白,1 380 838 440个相互作用。将丹参酮IIA治疗SCI的基因靶点导入String数据库,限定物种为人,获取蛋白相互作用关系,结果保存成TSV格式,导入Cytoscape3.6.0统计软件绘制相互作用网路。首先对此蛋白互作网络进行总体分析,然后使用Cytoscape3.6.0统计软件中的cytohubba插件确定最重要的5个蛋白互作关系,计算方法的选择最大团中心性(Maximal Clique Centrality,MCC)[11]。在网络中,用节点(Node)表示成分和靶点,用边(edge)表示成分与靶点之间的相互作用。
1.5 GO分析与KEGG通路分析 Metascape数据库(http://metascape.org)可为大规模的基因或蛋白提供系统综合的生物功能注释信息,能够找出最显著富集的生物学注释[12]。将将丹参酮IIA作用于SCI的基因靶点导入Metascape数据库,Input as species和Analysis as species均设置为H.sapiens,选择自定义分析(Custom Analysis)中的富集(Enrichment),分别对GO Biological Processes、GO Molecular Functions、GO Cellular Components、KEGG Pathway进行富集分析(Enrichment Analysis),保存结果,筛选P值排名靠前条目。GO分析用GraphPad Prism 7.0统计软件绘制条形图,KEGG Pathway,用R语言绘制气泡图。
2 结果
2.1 丹参酮IIA基因靶点预测 丹参酮IIA在TCMSP数据库中收录靶点44个,在BATMAN-TCM数据库中收录靶点27个,在DTPS数据库中收录靶点23个,在STITCHI数据库中收录靶点8个,在Swiss Target Prediction数据库收录靶点20个,通过文献挖掘收集靶点119个[13],将收集到的靶点汇总删除重复项后,得到丹参酮IIA相关靶点182个。
2.2 SCI相关基因靶点筛选 通过GeneCards、CTD和DisGeNET数据库以spinal cord injuries为关键词检索与其相关基因,分别得到4 428个、14 180个、8个结果,汇总删除重复基因后得到13 708个与SCI相关基因。
2.3 丹参酮IIA治疗SCI相关靶点的收集 将丹参酮IIA作用靶点与SCI相关发病机制靶点基因进行映射比较,发现157个共同基因,推测这些基因可能为丹参酮IIA治疗SCI的相关基因靶点。见图1。
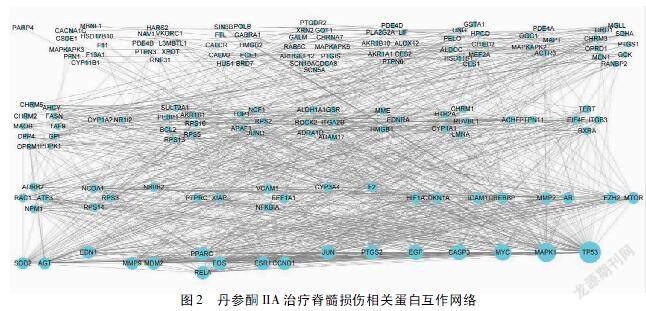
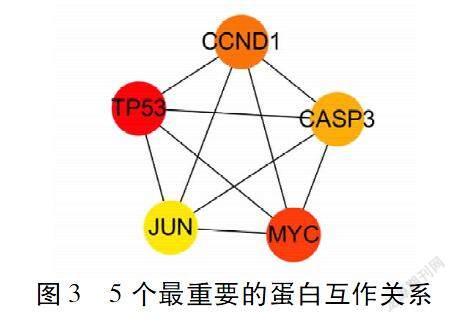
2.4 丹参酮IIA治疗SCI相关蛋白互作网络的构建与分析 丹参酮IIA治疗SCI的靶点蛋白互作网络结果详见图2,图中节点表示蛋白,边表示蛋白之间的关联,共涉及155个节点,1 129条边(其中VPS4、MMEL与其他蛋白没有相互作用,这2个蛋白不在相互作用网络中体现)。使用cytohubba插件中MCC方法计算后,确定最重要的5个蛋白互作关系(见图3),结果表明肿瘤蛋白p53(TP53)、Myc原癌基因蛋白(MYC)、G1/S-特异性周期蛋白-D1(CCND1)、半胱氨酸天冬氨酸蛋白酶3(CASP3)、JUN原癌基因蛋白(JUN)可能是此网络中较为重要的蛋白靶点。
2.5 GO分析 使用Metascape数据库对丹参酮IIA治疗SCI的相关靶点进行GO分析,结果BP分析(图4)中排名靠前的有药物反应(23个靶点/14.65%)、脂多糖反应(16个靶点/10.19%)、细胞对缺氧的反应(12个靶点/7.64%)、应对缺氧(14个靶点/8.92%)、细胞对DNA损伤刺激的反应(15个靶点/9.55%)等生物学过程。MF分析(图5)排名靠前的的有酶结合(28个靶点/17.83%)、转录因子结合(20个靶点/12.74%)、蛋白结合(122个靶点/77.71%)、药物结合(10个靶点/6.37%)、氧化还原酶活性(13个靶点/8.28%)等分子功能。CC分析(图6)排名靠前的有细胞质基质(72个靶点/45.86%)、核质(60个靶点/38.22%)、胞外外泌体(56个靶点/35.67%)、黏着斑(20个靶点/12.74%)、核染色质(13个靶点/8.28%)等细胞组分。
2.6 KEGG通路分析 使用Metascape数据库对行气活血通络方治疗神经根型颈椎病的157个靶点进行KEGG通路分析,根据P值筛选靠前的15个代表性通路见图7。图中节点的颜色从绿到红反应了P值从大到小,节点从小到大反应了相关基因数量的从少到多。进一步分析发现与SCI相关度最高的有6条通路,包括cAMP signaling pathway、TNF signaling pathway、HIF-1 signaling pathway、PI3K-Akt signaling pathway、MAPK signaling pathway、Apoptosis等信号通路。
3 讨论
脊髓损伤的治疗一直以来是医学界的难题,组织工程技术、干细胞技术治疗脊髓损伤都进行了大量的努力与尝试,但取得的进展却有限。成体中枢神经系统神经元的再生不是一个一步到位的过程,而是需要保护神经元存活,帮助其向原始神经元目标定向迁移扩展,重新髓鞘化并形成功能性突触的过程。因此,大量的理化事件同时发生才可能恢复受损神经元的功能[14]。近年来,在这一领域出现了联合治疗的趋势,以协同和提高单一治疗的疗效[15]。丹参酮IIA常用于治疗心脑血管疾病,也学者发现丹参酮IIA可以通过减轻炎性反应,减少细胞凋亡,逆转氧化还原不平衡状态等方面对受伤脊髓产生保护作用[4],但具体分子机制仍需更加深入的探索。
本研究通过网络药理学相关理念和技术,以多个数据库为背景结合文献检索,预测出丹参酮IIA拥有相关作用靶点182个,此种方法能更全面的预测药物单体的作用靶点,避免单一数据库的片面性和局限性。通过GeneCards、CTD和DisGeNET数据库检索得到13 708个与SCI相关基因。将丹参酮IIA作用靶点与SCI相关发病机制靶点进行映射比较,发现157基因可能为丹参酮IIA治疗SCI的相关基因靶点。将丹参酮IIA作用于SCI的靶点蛋白进行网络互作网分析发现,各靶点间存在大量的互作关系,而TP53、MYC、CCND1、CASP3、JUN可能为此网络中最重要的5个靶点蛋白。其中转录因子TP53参与DNA修复、细胞周期、细胞存活和轴突生长等过程。Floriddia E等人的研究发现,小鼠脊髓半切模型中,TP53基因敲除的小鼠与野生型小鼠比较运动功能恢复减慢,损伤区域瘢痕增生明显;且TP53基因缺失导致轴突断端生长锥发芽能力降低,并有明显的顶梢枯死现象。提示TP53是影响脊髓损伤后轴突功能恢复和胶质反应的关键修饰因子[16]。Myc与CCND1基因都属于细胞周期调控相关的基因,Di Giovanni S等人的研究表明,脊髓损伤后的4和24 h Myc、CCND1都有明显的过表达,继发引起炎性反应、氧化应激、DNA损伤等相关反应,最终导致神经元凋亡[17]。Zhao X等人发现兔脊髓缺血再灌注损伤后8 h CCND1和Cdk4蛋白的免疫反应在运动神经元核內被选择性诱,最终导致细胞凋亡,因此推测细胞周期蛋白CCND1和Cdk4的诱导可能与兔脊髓缺血后程序性细胞死亡的改变有关[18]。CASP3是细胞凋亡过程中最主要的终末剪切酶,在细胞凋亡中起着不可替代的作用。脊髓损伤后内质网应激可引起运动功能障碍、神经元凋亡、组织损伤和PI3K/AKT通路抑制。Li H等人的研究表明,淫羊藿苷显著抑制CASP3等内质网应激凋亡蛋白的表达,对脊髓损伤后运动恢复和神经元存活的神经保护作用[19]。JUN是一种核内原癌基因,被认为对神经再生过程起到重要作用。Danzi MC等检测不同含JUN的二聚体对背根神经节、皮质和海马神经元轴突生长的影响,结果表明,JUN和转录激活因子3(Atf3)组成的二聚体促进了大鼠中枢神经系统神经元和小鼠背根神经节神经元的轴突生长[20]。
GO富集分析发现,丹参酮IIA治疗SCI主要涉及药物反应、脂多糖反应、细胞对缺氧的反应、细胞对DNA损伤刺激的反应等生物学过程;涉及激酶结合、转录因子结合、蛋白结合、药物结合、氧化还原酶活性等分子功能;涉及细胞质基质、核质、细胞外泌体、黏着斑、核染色质等细胞组分。
通過KEGG生物途径富集分析发现,有15条信号通路被显著富集,其中环磷酸腺苷(cAMP)、丝裂原活化蛋白激酶(MAPK)、磷脂酰肌醇3激酶-蛋白激酶B(PI3K-AKT)、肿瘤坏死因子(TNF)、缺氧诱导因子1(HIF-1)、细胞凋亡(Apoptosis)等6条通路在丹参酮IIA治疗脊髓损伤过程中发挥着较为关键的作用。在胚胎中枢神经系统中,cAMP水平很高,同时轴突生长非常旺盛,出生后不久,背根神经节(DRG)神经元的cAMP水平迅速下降,其中枢神经轴突的再生能力也下降[21-22]。神经营养因子可通过增加cAMP水平,促进轴突生长和再生增加[23-24]。而在体内注射cAMP类似物(db-camp),也可激活下游效应器蛋白激酶A(PKA)和环磷腺苷效应元件结合蛋白(CREB),并促进脊髓中央支的再生[25]。MAPK在神经系统中发挥着多种作用,包括促进细胞增殖、神经可塑性和细胞存活。目前已知的MAPK有3类:细胞外信号调节激酶(Erks)、丝裂原活化蛋白激酶p38(p38 MAPK)和c-jun n-末端激酶(JNKs)。这些都可以进一步分为不同的子类,包括Erk 1/2亚型和不同的JNK蛋白,JNK 1、2和3,许多研究表明,这些通路可参与脊髓损伤的修复[26-27]。PI3K信号通常由细胞外生长因子激活受体酪氨酸激酶(RTK)或g蛋白偶联受体(GPCR)触发。一旦激活,PI3K可以磷酸化磷脂酰肌醇-4,5-磷酸(PIP2)形成PIP3。PIP3是一种多用途的二级信使,通过3-磷酸肌醇依赖蛋白激酶的活性(PDKs),促进蛋白激酶B(Akt)的活化及其膜定位[29]。SCI后病变区域内Akt磷酸化减少,而促进损伤区域PI3K增加可使磷酸化Akt增加从而增加神经元数量[30-31]。TNF作为一种重要的细胞因子,可诱导多种细胞内信号通路,包括凋亡、细胞存活、炎性反应和免疫。Karova K等研究表明,神经前体细胞移植可明显减轻受伤脊髓中TNF-α的产生,并且接受移植大鼠胶质瘢痕形成减少,灰质保存增强,空洞缩小[32]。HIF-1是一种转录因子,具有调节氧稳态的功能,它由HIF-1α和HIF-1β 2个亚基组成,HIF-1是许多缺氧诱导基因在低氧条件下的主调控因子。HIF-1的靶基因编码蛋白质,增加氧分子的传递,介导对氧分子剥夺的适应性反应。Siddiq A等的研究表明,缺氧诱导因子脯氨酰羟化酶1(HIF-PH1)的选择性抑制可通过HIF-和CREB-介导神经保护作用,抵抗常氧氧化死亡[33]。Apoptosis是一种通过活化天冬氨酸特异性半胱氨酸蛋白酶(caspases)来消除受损或冗余细胞的基因程序化过程,如何抑制脊髓损伤后神经元凋亡,从而促进神经功能恢复,减轻损伤程度是中枢神经领域研究的热点[34-36]。
综上所述,本研究利用网络药理学方法对丹参酮IIA治疗脊髓损伤的机制进行探讨,发现丹参酮IIA可能从多种信号通路和生物过程对脊髓损伤发挥治疗作用,也为中药单体治疗SCI提供了机制研究的思路。本研究的局限性在于仅基于网络药理学层面对丹参酮IIA治疗SCI进行了初探,在各数据库资料的准确性及数据更新等方面问题具有一定的局限性。基于本文得到的丹参酮IIA治疗SCI的相关通路及靶点的验证正在进行中。
参考文献
[1]Chhabra HS,Sarda K.Clinical translation of stem cell based interventions for spinal cord injury-Are we there yet?[J].Adv Drug Deliv Rev,2017,120:41-49.
[2]James S L,Theadom A,Ellenbogen R G,et al.Global,regional,and national burden of traumatic brain injury and spinal cord injury,1990-2016:a systematic analysis for the Global Burden of Disease Study 2016[J].The Lancet Neurology,2019,18(1):56-87.
[3]Eckert M J,Martin M J.Trauma:spinal cord injury[J].Surgical Clinics,2017,97(5):1031-1045.
[4]Yin X,Yin Y,Cao FL,et al.Tanshinone IIA attenuates the inflammatory response and apoptosis after traumatic injury of the spinal cord in adult rats[J].PLoS One,2012,7(6):e38381.
[5]Gong G,Gu Y,Zhang Y,et al.Tanshinone IIA alleviates oxidative damage after spinal cord injury in vitro and in vivo through up-regulating miR-124[J].Life Sci,2019,216:147-155.
[6]Zhang XM,Ma J,Sun Y,et al.Tanshinone IIA promotes the differentiation of bone marrow mesenchymal stem cells into neuronal-like cells in a spinal cord injury model[J].J Transl Med,2018,16(1):193.
[7]Yang YD,Yu X,Wang XM,et al.Tanshinone IIA improves functional recovery in spinal cord injury-induced lower urinary tract dysfunction[J].Neural Regen Res,2017,12(2):267-275.
[8]賀丰,穆晓红,付玲玲,等.脊髓损伤的中医研究现状[J].世界中西医结合杂志,2017,12(3):440-444.
[9]Hopkins A L.Network pharmacology[J].Nature Biotechnology,2007,25(10):1110-1111.
[10]von MC,Jensen LJ,Snel B,et al.STRING:known and predicted protein-protein associations,integrated and transferred across organisms[J].Nucleic Acids Res,2005,33(Database issue):D433-437.
[11]Meghanathan N.Maximal clique size versus centrality:a correlation analysis for complex real-world network graphs[C]Proceedings of 3rd International Conference on Advanced Computing,Networking and Informatics.Springer,New Delhi,2016:95-101.
[12]Tripathi S,Pohl MO,Zhou Y,et al.Meta-and Orthogonal Integration of Influenza “OMICs” Data Defines a Role for UBR4 in Virus Budding[J].Cell Host Microbe,2015,18(6):723-735.
[13]徐文华,郑景辉,赵阳,等.基于网络药理学和生物信息学的丹参酮ⅡA治疗冠心病的分子生物学机制分析[J].中草药,2019,50(5):1131-1140.
[14]Kim HY,Kumar H,Jo MJ,et al.Therapeutic Efficacy-Potentiated and Diseased Organ-Targeting Nanovesicles Derived from Mesenchymal Stem Cells for Spinal Cord Injury Treatment[J].Nano Lett,2018,18(8):4965-4975.
[15]Ruzicka J,Urdzikova LM,Kloudova A,et al.Anti-inflammatory compound curcumin and mesenchymal stem cells in the treatment of spinal cord injury in rats[J].Acta Neurobiol Exp(Wars),2018,78(4):358-374.
[16]Floriddia E,Nguyen T,Di GS.Chromatin immunoprecipitation from dorsal root ganglia tissue following axonal injury[J].J Vis Exp,2011,7(53):2803.
[17]Di GS,Knoblach SM,Brandoli C,et al.Gene profiling in spinal cord injury shows role of cell cycle in neuronal death[J].Ann Neurol,2003,53(4):454-468.
[18]Zhao X,Wang XW,Zhou KS,et al.Expression of Ski and its role in astrocyte proliferation and migration[J].Neuroscience,2017,362:1-12.
[19]Li H,Zhang X,Qi X,et al.Icariin Inhibits Endoplasmic Reticulum Stress-induced Neuronal Apoptosis after Spinal Cord Injury through Modulating the PI3K/AKT Signaling Pathway[J].Int J Biol Sci,2019,15(2):277-286.
[20]Danzi MC,Mehta ST,Dulla K,et al.The effect of Jun dimerization on neurite outgrowth and motif binding[J].Mol Cell Neurosci,2018,92:114-127.
[21]Cai D,Qiu J,Cao Z,et al.Neuronal cyclic AMP controls the developmental loss in ability of axons to regenerate[J].J Neurosci,2001,21(13):4731-4739.
[22]Chaudhry N,Filbin MT.Myelin-associated inhibitory signaling and strategies to overcome inhibition[J].J Cereb Blood Flow Metab,2007,27(6):1096-1107.
[23]Gao Y,Nikulina E,Mellado W,et al.Neurotrophins elevate cAMP to reach a threshold required to overcome inhibition by MAG through extracellular signal-regulated kinase-dependent inhibition of phosphodiesterase[J].J Neurosci,2003,23(37):11770-11777.
[24]Lu P,Yang H,Jones LL,et al.Combinatorial therapy with neurotrophins and cAMP promotes axonal regeneration beyond sites of spinal cord injury[J].J Neurosci,2004,24(28):6402-6409.
[25]Neumann S,Bradke F,Tessier-Lavigne M,et al.Regeneration of sensory axons within the injured spinal cord induced by intraganglionic cAMP elevation[J].Neuron,2002,34(6):885-893.
[26]Lee JY,Chung H,Yoo YS,et al.Inhibition of apoptotic cell death by ghrelin improves functional recovery after spinal cord injury[J].Endocrinology,2010,151(8):3815-3826.
[27]Wang HY,Crupi D,Liu J,et al.Repetitive transcranial magnetic stimulation enhances BDNF-TrkB signaling in both brain and lymphocyte[J].J Neurosci,2011,31(30):11044-11054.
[28]Tran HT,Sanchez L,Brody DL.Inhibition of JNK by a peptide inhibitor reduces traumatic brain injury-induced tauopathy in transgenic mice[J].J Neuropathol Exp Neurol,2012,71(2):116-129.
[29]Ghasemlou N,Lopez-Vales R,Lachance C,et al.Mitogen-activated protein kinase-activated protein kinase 2(MK2)contributes to secondary damage after spinal cord injury[J].J Neurosci,2010,30(41):13750-13759.
[30]Walker CL,Walker MJ,Liu NK,et al.Systemic bisperoxovanadium activates Akt/mTOR,reduces autophagy,and enhances recovery following cervical spinal cord injury[J].PLoS One,2012,7(1):e30012.
[31]Howitt J,Lackovic J,Low LH,et al.Ndfip1 regulates nuclear Pten import in vivo to promote neuronal survival following cerebral ischemia[J].J Cell Biol,2012,196(1):29-36.
[32]Karova K,Wainwright JV,Machova-Urdzikova L,et al.Transplantation of neural precursors generated from spinal progenitor cells reduces inflammation in spinal cord injury via NF-κB pathway inhibition[J].J Neuroinflammation,2019,16(1):12.
[33]Siddiq A,Aminova LR,Troy CM,et al.Selective inhibition of hypoxia-inducible factor(HIF)prolyl-hydroxylase 1 mediates neuroprotection against normoxic oxidative death via HIF-and CREB-independent pathways[J].J Neurosci,2009,29(27):8828-8838.
[34]Seo JY,Kim YH,Kim JW,et al.Effects of Therapeutic Hypothermia on Apoptosis and Autophagy After Spinal Cord Injury in Rats[J].Spine(Phila Pa 1976),2015,40(12):883-890.
[35]Gensel JC,Zhang B.Macrophage activation and its role in repair and pathology after spinal cord injury[J].Brain Res,2015,1619:1-11.
[36]Brotfain E,Gruenbaum SE,Boyko M,et al.Neuroprotection by Estrogen and Progesterone in Traumatic Brain Injury and Spinal Cord Injury[J].Curr Neuropharmacol,2016,14(6):641-653.
(2019-06-22收稿 責任编辑:王明)
基金项目:国家自然科学基金项目(81774330);中国博士后科学基金项目(2018M641608)作者简介:贺丰(1986.09—),男,博士,主治医师,研究方向:脊柱脊髓相关疾病研究,Tel:(010)84734120,E-mail:dr_hefeng@sina.com通信作者:朱立国(1961.09—),男,博士,教授,主任医师,研究方向:脊柱脊髓相关疾病的研究,Tel:(010)84734120,E-mail:zhlg95@aliyun.com
