盐胁迫对鲁氏酵母菌生理特性的影响
2019-08-27王定康周荣清吴重德
王定康, 张 敏, 黄 钧, 金 垚, 周荣清, 吴重德
(四川大学 轻工科学与工程学院/皮革化学与工程教育部重点实验室,四川 成都 610065)
鲁氏酵母菌(Zygosaccharomycesrouxii)广泛存在于传统发酵食品如豆瓣、酱油等生产过程中,在产品产醇、产香、增鲜等方面具有重要作用[1-2]。在食品发酵过程中,鲁氏酵母菌可以产生风味物质,如异戊醇、异丁醇、活性戊醇等重要的芳香杂醇类物质,以提高食品品质[3]。然而,在发酵食品生产过程中,鲁氏酵母菌经常受到多种环境胁迫,包括盐胁迫、酸性胁迫和酒精胁迫等,特别是盐胁迫引起的渗透胁迫会导致细胞结构和生理的损伤[4];同时盐胁迫也会引起细胞内活性氧(ROS)的形成,从而影响胞内氧化还原稳态的变化,而维持氧化还原稳态被认为是细胞耐盐性能的重要因素之一[5]。研究表明,细胞会采取一些策略来抵御盐胁迫,比如调节细胞内微环境、细胞膜和细胞代谢等[6]。He等[7]研究了嗜盐四链球菌盐胁迫耐受机制,发现盐胁迫使细胞内脯氨酸、甘氨酸甜菜碱和海藻糖含量升高。Sara等[8]通过补充油酸和麦角固醇,发现可以降低酿酒酵母胞内活性氧的含量,从而降低对细胞膜的氧化损伤。谷胱甘肽(GSH)是一种由谷氨酸、半胱氨酸和甘氨酸组成的三肽,在生物系统中具有抗氧化、增强免疫、细胞解毒活性和营养调节等作用[9]。Zhang等[10]研究发现,GSH可以通过调节细胞生理功能,提高旧金山乳杆菌的抗冷冻胁迫性能。
近些年,对鲁氏酵母菌耐盐机制尚未充分研究,为发挥食品微生物鲁氏酵母菌的生理功能和提高其生物制造效率,有必要深入认识鲁氏酵母菌盐胁迫耐受的生理机制。本文拟研究盐胁迫对鲁氏酵母菌生理特性的影响,希望为进一步认识鲁氏酵母菌的耐盐机制,提升其盐胁迫抗性,强化其发酵生产性能奠定理论基础。
1 材料与方法
1.1 材料与试剂
1.1.1菌株
鲁氏酵母菌(Zygosaccharomycesrouxii),由本实验室从百年老字号酿造企业的酱醪中分离,经26S rDNA和ITS区域序列测序、鉴定、命名,并保藏于中国普通微生物菌种保藏中心,保藏编号为CGMCC NO. 3791。
1.1.2实验试剂
HEPES-K细胞裂解液,购自雷根生物公司;BCECF AM荧光探针,购自北京百奥莱博科技有限公司;过氧化氢酶CAT测定试剂盒、超氧化物歧化酶SOD测定试剂盒、过氧化物酶POD测定试剂盒、谷胱甘肽过氧化物酶GSH-PX测定试剂盒,均购自南京建成生物公司;蛋白浓度测定试剂盒,购自碧云天生物技术研究所。其他试剂均为国产分析纯。
1.2 仪器与设备
GL-20G-Ⅱ型高速冷冻离心机,上海安亨科学仪器厂;92-IIN 型超声波破碎粉碎仪,宁波新芝生物科技股份有限公司;TU-1901型双光束紫外可见分光光度计,上海沪粤明科学仪器有限公司;SW-CJ-2G型超净工作台,苏州净化设备有限公司;JJ500型分析天平,常熟双杰测试仪器厂;F-7100型荧光分光光度计,日本日立公司。
1.3 实验方法
1.3.1培养基及盐胁迫实验
YPD培养基:10 g/L酵母膏,20 g/L蛋白胨,20 g/L 葡萄糖,pH值为6.0。
从甘油管中将菌种Z.rouxii以体积分数为5%的接种量接种于新鲜的YPD培养基中,30 ℃静置培养24 h,至对数中期获得种子液。将体积分数为5%的种子液接种于不同NaCl质量浓度(0、60、120 g/L)的新鲜YPD培养基中,30 ℃静置培养36 h,至对数中后期,进行后续生理数据测定分析。
1.3.2鲁氏酵母菌生物量和乙醇含量测定
菌体生物量的测定:取不同盐浓度下培养至对数中后期的细胞,10 000g离心5 min,用蒸馏水洗菌两次,105 ℃烘干2 h,测定干细胞重量。
乙醇含量测定:参考文献[11]所述的方法与步骤。
1.3.3胞内pH值测定
参考文献[12]所述的方法,取5 mL 生长至对数中后期的细胞, 10 000g离心5 min,用50 mmol/L HEPES-K (pH值8.0)缓冲液洗涤2次,重悬于等体积的相同缓冲液,加入1 μL BCECF AM,30 ℃温水浴20 min。用50 mmol/L磷酸缓冲液(pH值7.0)洗涤3次后重悬于等体积的该缓冲液中。根据文献[13-14]所述方法用荧光分光光度计测定菌悬液荧光强度(Itotal)和上清液荧光强度(Ifiltrate),其中激发波长为488 nm和440 nm,发射波长为535 nm,狭缝宽度均为5 nm。荧光强度计算公式见式(1)。
I=[(I490)total-(I490)flitrate]/[(I440)total-(I440)flitrate],
(1)
再根据lgI的值由标准曲线计算胞内pH值。
1.3.4胞内活性氧水平及抗氧化酶系活力测定
取5 mL 生长至对数中后期的细胞, 10 000g离心5 min,收集菌体,用50 mmol/L磷酸缓冲液(pH值7.0)洗涤3次,重悬于等体积的该缓冲液中。将样品用超声细胞破碎仪破碎,破碎条件参照文献[15]的方法,破碎后10 000g离心5 min,取上清液。采用南京建成生物公司的活性氧(ROS)试剂盒测定胞内ROS含量,并用相关抗氧化酶试剂盒测定抗氧化酶酶活力,具体操作参照说明书进行,用紫外分光光度计检测其吸光值。
1.3.5胞内GSH含量测定
细胞提取和破碎方法同1.3.4,采用南京建成生物公司的还原型GSH试剂盒测定胞内GSH含量,具体操作参照说明书进行,用紫外分光光度计检测其吸光值。
1.4 数据处理
论文数据均为3次重复实验的平均值。采用软件SPSS 19.0对数据进行统计学分析,以 One-way ANOVA 进行显著性检验,并采用软件Origin 8.5对实验数据绘图。
2 结果与分析
2.1 盐胁迫对鲁氏酵母菌生长和代谢的影响
考察了不同盐浓度胁迫对鲁氏酵母菌生长和代谢的影响,结果如图1。鲁氏酵母菌在无盐培养条件下生长性能良好,细胞干重可达1.2 mg/mL。随着盐质量浓度增加至120 g/L,生物量逐渐下降,细胞干重降到0.81 mg/mL,表明盐胁迫会对鲁氏酵母菌的生长产生抑制作用,但该菌具有一定的耐盐性能。然而,随着盐质量浓度从0增加到120 g/L,发酵液中单位(每克)菌体的乙醇含量却逐渐升高,从11.64 mg增加到19.20 mg,这说明盐胁迫下鲁氏酵母菌单位菌体的乙醇含量增加,可能与盐胁迫后生成乙醇的代谢通量增强有关。李明达等[16]的研究也表明,盐胁迫下酿酒酵母合成乙醇代谢途径的碳流量增加了23.32%,可能是因为较高的盐浓度抑制了酵母的有氧呼吸作用,增加了厌氧发酵产物乙醇的通量,这与本研究中120 g/L盐质量浓度下乙醇含量显著增加的结果一致。
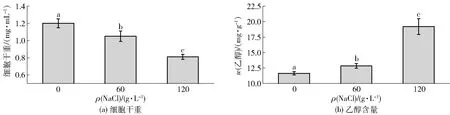
不同小写字母表示差异显著(P<0.05)(n=3)。图1 盐胁迫对鲁氏酵母菌细胞干重和乙醇含量的影响Fig.1 Changes of biomass and ethanol content in Z. rouxii under salt stress
2.2 盐胁迫对鲁氏酵母菌胞内pH值的影响
测定了鲁氏酵母菌盐胁迫后胞内pH水平的变化情况,见图2。图2显示,鲁氏酵母菌胞内pH值随着盐浓度的增加而降低,在120 g/L盐质量浓度下较为显著。据报道,细胞胞内pH值与细胞生长活力有关,pH值的减小伴随着细胞生长活力减弱[17],细胞在120 g/L盐质量浓度下,胞内pH值降低与其生长性能降低的结果相一致。赵春燕等[14]研究发现,乳酸链球菌素产生菌在酸胁迫条件下,胞内pH值水平降低,其生长也受到一定抑制。张娟[18]报道了冷冻胁迫会导致旧金山乳杆菌胞内pH水平降低,发现GSH能显著改善冷冻胁迫造成的胞内pH值的下降。

不同小写字母表示差异显著(P<0.05)(n=3)。图2 鲁氏酵母菌盐胁迫下胞内pH水平变化Fig.2 Changes of intracellular pH in Z. rouxii under salt stress
2.3 盐胁迫对鲁氏酵母菌胞内活性氧水平和抗氧化酶系活力的影响
活性氧(reactive oxygen species, ROS)是生物体中主要的氧自由基的统称,包括超氧阴离子自由基、羟自由基、脂氧自由基、二氧化氮和一氧化氮自由基,并且高浓度的活性氧簇对细胞机体有毒害作用[19]。本研究考察了盐胁迫对鲁氏酵母菌胞内活性氧ROS水平的影响(图3),结果表明,随着盐胁迫浓度增加,胞内活性氧ROS水平急剧增加,特别是在高盐浓度(120 g/L)时最为明显,比NaCl浓度为0时提高了21倍。说明高盐环境会同时引起细胞内氧胁迫,从而对细胞造成氧化损伤[20]。

不同小写字母表示差异显著(P<0.05)(n=3)。图3 鲁氏酵母菌盐胁迫下胞内ROS水平变化Fig.3 Changes of intracellular ROS level in Z. rouxii under salt stress
微生物细胞自身具有抵御ROS损伤的能力,如过量表达抗氧化酶系,包括过氧化氢酶(CAT),超氧化物歧化酶(SOD)、过氧化物酶(POD),以及GSH过氧化物酶(GSH-PX);此外,还有低分子量的抗氧化剂如GSH等都可以帮助机体抵御氧化损伤[21]。本研究考察了盐胁迫对鲁氏酵母菌抗氧化酶系活性的影响,实验结果如图4。测定了CAT、SOD、POD以及GSH-PX的活性,研究结果表明,鲁氏酵母菌的抗氧化酶系的活力在盐胁迫后均有显著性提高,在60 g/L NaCl时,POD酶活力提高,比未经盐胁迫的活性提高了1.8倍;继续提高盐质量浓度至120 g/L,酶活性略有降低。可能是细胞机体为了分解体内的活性氧,提高了抗氧化酶系的活力,这可能是Z.rouxii具有较高耐盐性能的原因之一[22]。Farhangi-Abriz等[23]报道了盐胁迫会导致胞内活性氧的过量产生,破坏细胞膜,而细胞的抗氧化酶系活性的提高在保护细胞免受氧化损伤方面发挥重要作用。此外,Jakubowski等[24]研究发现,随着酵母菌中抗氧化物质含量增加,比如超氧化物歧化酶SOD的活力增加等,酵母菌在进入稳定期后会对氧化损伤的增强做出适应性反应。
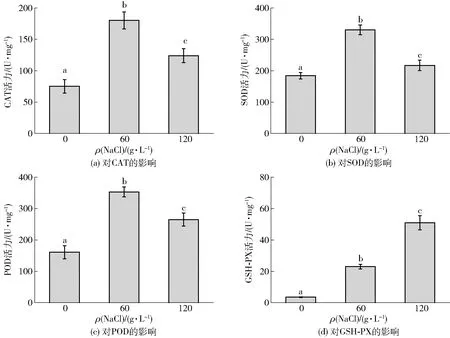
不同小写字母表示差异显著(P<0.05)(n=3)。图4 盐胁迫对鲁氏酵母菌胞内抗氧化酶活力的影响Fig.4 Effects of salt stress on intracellular antioxidant enzymes activity in Z. rouxii
2.4 盐胁迫对鲁氏酵母菌胞内GSH含量的影响
GSH有助于维持细胞内氧化还原稳态,保护细胞免受氧化损伤[25]。本研究考察了盐胁迫后鲁氏酵母菌细胞内GSH含量变化,实验结果见图5。图5显示,盐胁迫后鲁氏酵母菌胞内GSH的含量逐渐增加,在120 g/L盐质量浓度时,GSH含量显著增加,与无盐条件相比增加了73.4%,有助于鲁氏酵母菌维持胞内氧化还原反应,并通过酶促反应维持体内稳态,从而抵御盐胁迫[26]。相似的结果在对热带假丝酵母的研究中也有报道,Ilyas等[27]发现热带假丝酵母菌在重金属离子胁迫条件下,胞内GSH的含量显著高于非胁迫状态下的,而氧化应激引起的GSH积累的增加可能是细胞存活的一个重要因素。

不同小写字母表示差异显著(P<0.05)(n=3)。图5 鲁氏酵母菌盐胁迫下胞内GSH含量变化Fig.5 Changes of intracellular glutathione content in Z. rouxii under salt stress
2.5 添加GSH对鲁氏酵母菌生长性能的影响
为了进一步研究GSH对鲁氏酵母菌耐盐性能的影响,考察了不同盐浓度下外源添加GSH对鲁氏酵母菌生长性能的影响,结果如图6。图6显示,在低盐条件下,添加0.5 g/L GSH的鲁氏酵母菌与未添加相比,生物量无显著区别。随着盐浓度的提高,在120 g/L NaCl质量浓度条件下,添加0.5 g/L GSH使得鲁氏酵母菌的生物量提高了15%。这说明,添加GSH能够提高鲁氏酵母菌盐胁迫下的生长性能。相似的结果在其他微生物研究中也有报道,Zhang等[4]研究了乳酸乳球菌的耐盐性能,发现在5 mol/L盐浓度下,含GSH的乳酸乳球菌细胞存活率显著高于对照组细胞。Lee等[28]研究发现,唾液乳杆菌在添加GSH的低pH值培养基中,比对照组生长速度快,生长性能更强。

*表示差异显著(P<0.05)(n=3)。图6 外源添加GSH对鲁氏酵母菌生长性能的影响Fig.6 Effect of exogenous glutathione addition on growth of Z. rouxii under salt stress
3 结 论
研究了盐胁迫对鲁氏酵母菌(ZygosaccharomycesrouxiiCGMCC 3791)细胞生理特性的影响。分别考察了盐胁迫对鲁氏酵母菌生长代谢以及胞内微环境(胞内pH值、胞内活性氧和抗氧化酶系水平等)的影响。研究结果表明,盐胁迫抑制了鲁氏酵母菌生长,提高了单位菌体的乙醇含量;同时研究发现,盐胁迫导致胞内pH水平降低,胞内活性氧水平和抗氧化酶系活力和胞内GSH含量均显著升高,从而可帮助细胞抵御盐胁迫及盐胁迫引起的氧胁迫。此外,通过外源添加GSH,使鲁氏酵母菌在120 g/L盐质量浓度下生物量提高了15%。本研究结果有助于了解鲁氏酵母菌盐胁迫条件下的生理特性及盐胁迫响应机制,可为进一步提高鲁氏酵母菌的耐盐性能,提升其食品微生物制作的效率提供理论基础。
