超富集植物对稀土元素吸收转运解毒与分异的研究进展*
2019-08-24陈莺燕刘文深郭美娜HUOTHermine汤叶涛仇荣亮
陈莺燕 刘文深 袁 鸣 郭美娜 刘 畅 HUOT Hermine汤叶涛,2,3† 仇荣亮,2,3
(1 中山大学环境科学与工程学院,广州 510275)(2 广东省环境污染控制与修复技术重点实验室(中山大学),广州 510275)(3 广东省土壤重金属污染修复工程技术研究中心,广州 510275)
稀土元素(R E E s)是由1 5 种镧系元素镧(La)、铈(Ce)、镨(Pr)、钕(Nd)、钷(Pm)、钐(Sm)、铕(Eu)、钆(Gd)、铽(Tb)、镝(Dy)、钬(Ho)、铒(Er)、铥(Tm)、镱(Yb)、镥(Lu),及其性质相近的钇(Y)和钪(Sc)共同组成的一类化学性质相似的元素[1]。稀土作为世界上极为重要的战略资源,在现代高科技行业、清洁能源发展和农牧业生产中发挥着日益重要的作用[2-3]。但随着稀土需求量的与日俱增,稀土矿山开发加剧,产生了大面积的稀土废弃尾砂地和稀土污染农田[4-5]。同时稀土在环境中的大量蓄积,可能导致稀土通过食物链、水和呼吸等途径进入人体并在体内器官中分布[6],并对人体健康产生潜在危害[7]。据报道,高稀土浓度暴露地区儿童的智商(IQ)显著低于无污染地区儿童的智商[8];长期的稀土暴露也可能会对人体循环系统和免疫系统造成损害[9],降低人体神经系统的传导速度[10],增加动脉硬化的发生率[11]等。因此,自20世纪90年代以来稀土被列为中国主要化学污染物之一[12],近年在国际上也被认为是一类新型污染物[13-14]。另一方面,2010年“欧盟关键原材料报告”[15]和2011年美国能源局(DOE)“关键原材料战略”[16]均指出,稀土已成为最紧缺和供应风险最高的原材料。因此从低品位稀土矿和稀土污染土壤中实现稀土资源化再利用已成为当前国际上的研究热点[17-18]。
植物采矿为实现稀土污染地生态治理和稀土资源化提供了一种经济有效的解决途径。植物采矿是指在金属污染地上种植超富集植物,修复污染土壤、恢复植被的同时,还能通过收获植物地上部实现金属资源化。超富集植物的选择是植物采矿成功的关键因素。稀土超富集植物是指能够超量吸收稀土并将其运移到地上部的植物,且其具有极端稀土耐受性,在稀土污染土壤上能正常生长,不会出现稀土毒害现象[19]。目前判定植物是否超富集稀土元素需要满足两个条件:一是地上部稀土元素含量达到或超过1 000 μg·g-1的植物[19],这一判定标准与其他微量金属(C o、C u、N i、P b、S b)、类金属(A s)和非金属(S e)的判定标准相同,即超富集植物中金属含量通常较正常植物高2 或3 个数量级[20];二是植物地上部稀土富集系数(Bio-concentration f a c t o r,B F,植物地上部的稀土浓度与土壤中相应稀土浓度的比值)达到或超过1,表明其具有较强的稀土元素吸收转运能力[19,21]。目前国内外已发现稀土超富集植物及稀土富集植物2 0多种[22-31],主要分布在蕨类植物,以及胡桃科(Juglandaceae)、商陆科(Phytolaccaceae)和大戟科(Euphorbiaceae)等双子叶植物中。其中蕨类植物芒萁(Dicranopteris dicthotoma)是目前叶片中稀土积累浓度最高的植物,可达3 358 μg·g-1[22],其地上部选择富集轻稀土元素(light REEs,LREEs,包括Pr、Nd、Pm、Sm、Eu、Gd),轻稀土的富集系数(BF)及转运系数(Translocation factor,TF,植物地上部的稀土浓度与植物根系中相应稀土浓度的比值)均大于重稀土元素(heavy REEs, HREEs,包括Tb、Dy、Ho、Er、Tm、Yb、Lu、Y)(表1)[32];大生物量双子叶植物美洲商陆(Phytolacca americana L.)叶片稀土含量最高可达1 040 μg·g-1[23],其地上部选择富集HREE,HREE的富集系数(BF)及转运系数(TF)均大于LREE(表1)[23]。全面了解超富集植物对稀土的吸收转运机理是植物采矿的重要前提条件。但迄今为止,超富集植物对稀土的吸收转运机制的研究远远滞后于对Ni、Zn、As、Cd、Pb等重金属或类金属的研究。
稀土元素具有相似的物理化学性质,但其存在的细小差异导致稀土元素在地球化学过程中产生分异。稀土分异在过去几十年中已被广泛应用于地学领域;在土壤-植物系统中稀土元素也存在明显的分异,从而在指示稀土吸收-转运机理方面展示出很大的潜力[33]。因此,本文将根据超富集植物富集稀土的几个关键过程,如根际土壤溶液化学过程、根系吸收、木质部转运及地上分布解毒等,系统综述根际土壤-植物体系中稀土的迁移机制及其与分异的联系,并提出超富集植物中稀土分异的概念模型,最后展望稀土超富集植物修复应用研究的发展前景,以期为稀土污染土壤的植物修复提供理论参考。
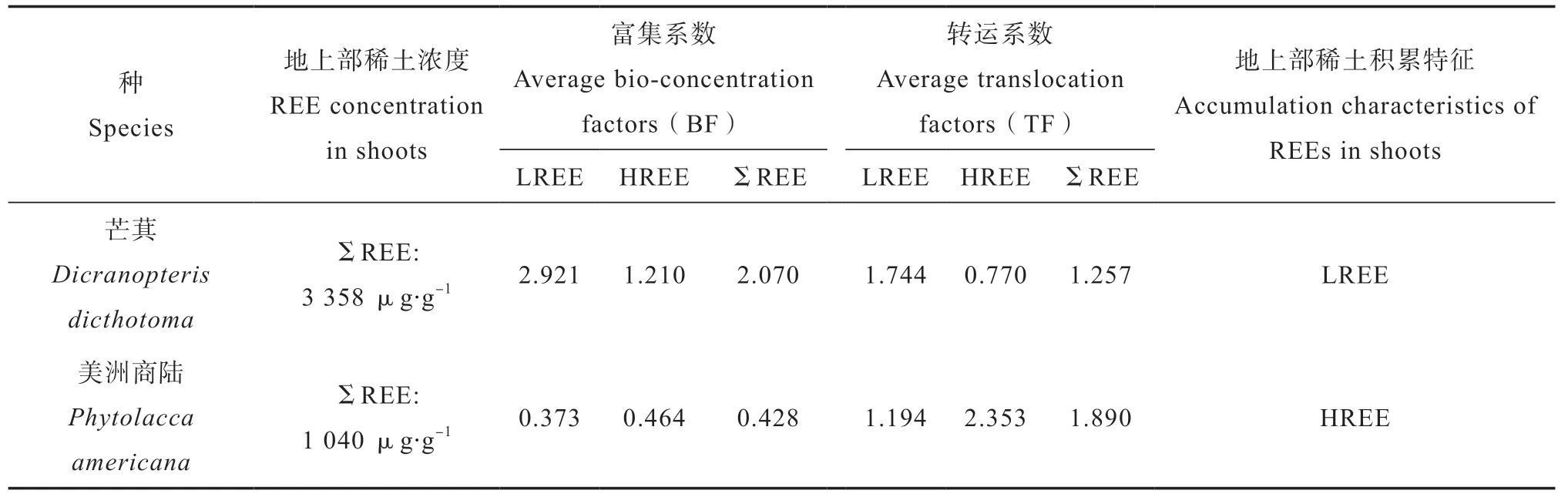
表1 稀土在代表性超富集植物地上部的积累特征[22-23,32]Table 1 Accumulation characteristics of REEs in shoots of representative hyperaccumulators
1 稀土的根际土壤溶液化学过程与分异
大多数情况下,植物根系主要吸收土壤溶液中的自由离子态稀土(free REE3+)。对超富集植物而言,由于其吸收速率较快,当自由离子态金属供应不足时,土壤溶液中其他形态的金属转化为可利用态金属供超富集植物吸收。Luo等[34]研究表明,Ni超富集植物在低/高有机质两种土壤中,自由离子态Ni的扩散过程均受到抑制。Wei等[35]发现,自然条件下生长的稀土超富集植物芒萁根系中稀土的分异模式与土壤A层中有效态稀土(1.0 mol·L-1NaOAc-HOAc,pH=4.8)的分异模式非常相似,认为芒萁根系主要吸收土壤中自由离子态稀土;芒萁根系与土壤A层中有效态稀土均出现Ce负异常现象,进一步表明土壤溶液中稀土的含量及其存在形态很大程度上决定了植物对稀土的吸收。
稀土的自由离子态浓度和其生物有效性受根际过程影响明显[36]。如植物根系在生长过程中可分泌草酸、柠檬酸等小分子有机酸,促使稀土从土壤颗粒表面解吸,继而与稀土生成络合物从而影响稀土离子向根表的扩散[37]。向土壤中添加酸醋和苹果酸大幅提高小麦根系对La的吸收[38]以及地上部La的含量[39];添加小分子有机酸和氨基酸促进超富集植物芒萁根系对稀土的吸收[40];向根系溶液中添加有机配体EDTA(乙二胺四乙酸)、柠檬酸和DTPA(二乙基三胺五乙酸),小麦根和叶中轻稀土轻微富集,重稀土相对亏损[41]。表明添加有机配体改变了根系对稀土的选择性吸收过程,进而影响其向上转运。此外,当自由离子扩散成为速控步骤时,金属络合物在根表重新解离,增加局部的自由离子浓度,从而促进金属的吸收。该方式主要发生在金属浓度极低(nmol级别)或扩散速率较小的环境介质[42],或吸收界面较大的生物[43]。由此可见,根际有机配体的种类和浓度是决定植物中稀土生物有效性及其分异模式的重要因素。
此外,根际土壤溶液中pH、Eh和无机离子含量等也能通过改变根际环境影响根际稀土的生物有效性,进而影响植物中稀土的含量和分异。如研究发现低pH时土壤中轻重稀土均易被溶解出来,使小麦对稀土的吸收选择性降低,轻重稀土之间的分异较小;而高pH时重稀土更易沉淀,小麦优先选择吸收轻稀土[44]。pH或Eh的降低有利于红壤中吸附在Fe-Mn氧化物上的La和Ce解离释放,使交换态和Fe-Mn氧化物结合态稀土减少[45],从而促进植物吸收稀土。土壤中Fe氧化物含量显著影响脐橙果实中稀土的浓度,可能是由于土壤中的稀土来源于溶解的羟基氧化铁[46]。以上因素的复合作用影响了稀土离子向根表的扩散,从而使稀土在植物体内的分布模式产生差异。
2 植物根系对稀土的吸收与分异
当土壤溶液中的稀土离子扩散至根表,根表细胞壁上带羧基或羟基的纤维素、果胶等大分子物质会通过吸附、沉淀、络合等作用结合根表质外体上的稀土,减少其进入细胞原生质体中[39]。如小麦根中约60%的稀土与细胞壁结合,其中果胶结合态稀土比例达88%[41]。XAS分析表明,小麦根细胞壁吸附主要发生在酸性较强(pH=3)的溶液体系,当pH升高至5.0时,稀土离子主要以磷酸盐沉淀的形式存在[47]。这两种机制的共同作用,导致根部产生中稀土富集和四重效应等分异特征[47]。Shan等[40]通过电镜研究,在芒萁根部内皮层和中柱细胞的细胞壁、细胞间质、细胞质膜和液泡中均检测到轻稀土沉积,表明稀土在根部的运输通过质外体和共质体途径共同完成。但目前质外体与共质体途径在超富集植物稀土吸收过程的作用与贡献尚未明确。理论上推测,因为将稀土固持在根细胞壁对其向地上转运不利,质外体吸附作用应该不是稀土超富集的主要机制。但目前关于稀土如何通过共质体途径被超富集植物根系吸收还鲜有报道。由于稀土尚未被证实为植物必需元素,植物不太可能演化出稀土的专性吸收系统,推测稀土很可能通过其他元素的跨膜运输通道进入根部原生质体。
2.1 被动扩散
稀土离子(9.6~11.5 nm)与Ca2+(9.9 nm)具有相似的离子半径,在细胞中通常与Ca2+结合在相同的位置上,表现出相似的行为特征。如在水稻[48]和黑麦[49]中发现Ca2+与La3+、Ce3+或Eu3+等具有拮抗作用;在植物根细胞膜中La3+被用作Ca2+通道阻断剂[50];Y3+、Ce3+等与Ca2+竞争细胞质膜上的转运蛋白,影响Ca2+的吸收转运[51];因此推测稀土可能通过Ca2+通道进入植物中。对稀土超富集植物的研究发现,野外芒萁植物整体相对于土壤介质更易富集轻稀土而耗损重稀土[40,52-53],与稀土分异理论认为扩散等动力学反应优先选择轻稀土的观点一致,意味着离子通道扩散可能在芒萁吸收稀土过程中起主导作用。此外,轻稀土离子半径与Ca2+更为相近,较重稀土更倾向于使用Ca2+通道进入根系[54]。因此推测通过Ca2+通道被动扩散进入根细胞是芒萁根部富集轻稀土的主要机制。水培研究也发现,美洲商陆整体相对溶液也富集轻稀土[54],与芒萁类似,说明Ca2+通道扩散可能是轻稀土进入美洲商陆根部的一种重要途径。
2.2 主动吸收
除了Ca2+通道,很多研究还报道植物根系对稀土的吸收可能与Al、Fe和Si等元素有关。例如,研究表明La可以抑制黑麦木质部汁液中Al含量[55],大幅提高水稻Al转运子Nrat1的表达[56]。在Rhynchanthera grandiflora叶片中也发现Al和La含量存在高度相关性[57],说明稀土可能通过Al的吸收通道进入植物中。水培实验表明,施加能量代谢抑制剂显著抑制美洲商陆根系稀土含量,说明其对稀土的吸收存在一个主动的、消耗能量的过程,同时Al3+的处理抑制了美洲商陆对稀土尤其是对重稀土的吸收,根中轻稀土与重稀土的比值(ΣLREE/ΣHREE)显著增加,表明美洲商陆根系中重稀土可能与Al竞争相同的吸收通道[54]。因此推测重稀土以自由离子态形式通过Al转运蛋白主动吸收进入根部也是美洲商陆吸收稀土的一种途径。美洲商陆根部相对轻稀土富集和重稀土亏损,一方面可能是由于在稀土吸收中Ca2+通道起着较Al转运蛋白更重要的作用,另一方面可能是由于从根部向地上部转运的过程中,重稀土较轻稀土有更高的转运能力[54]。
稀土也可能通过铁通道蛋白被根系吸收。研究表明,Dy3+、Er3+、Tm3+和Lu3+能与铁竞争转铁蛋白(transferrin)上的结合位点,与转铁蛋白受体结合形成复合物进入细胞内[58]。野外调查也显示,山毛榉、云杉、橡树根系稀土的吸收与铁具有相同的吸收模式,表明稀土可能通过铁通道蛋白跨膜运输[59]。稀土还被报道与Si存在相关性。例如富硅植物赤竹根部Si的分布与土壤硅酸盐组分中的稀土分异规律一致[60];在荚果蕨类根部皮层中发现与土壤中硅酸盐矿物组成相似的含稀土硅石颗粒,说明该植物吸收的稀土来自土壤中的硅矿物[61]。然而,植物是否能够直接吸收稀土与硅形成的螯合物仍然需要进一步的证据。
由此可见,超富集植物根系稀土吸收不仅涉及质外体吸附和共质体运输途径,还可能存在多种不同的跨膜运输系统,因此有必要开展根系吸收动力学、生物化学和植物生理学的研究,结合稀土形态与分异特征分析以揭示超富集植物根系对稀土的高效吸收策略。
3 稀土在木质部的转运与分异
稀土进入共质体和质外体后跨过凯氏带,然后经过植物的木质部向地上部转运。在植物木质部中,稀土通常以自由离子态或有机络合态形式向地上部转运。稀土的转运形态不同,使该过程在不同植物中产生完全不同的稀土分异模式。
研究显示,Ni、Zn等重金属在植物木质部中可能主要以离子态转运[62-63]。类似地,稀土也可能以自由离子态在植物木质部转运。野外研究显示,芒萁从根部向上运输过程不断富集轻稀土,尤其茎(La/Yb=24,La/Yb: 归一化镧镱比,描述轻重稀土元素之间的分异特征)与叶柄(La/Yb=136)之间分异最为显著,意味着木质部中稀土可能主要以自由离子态随蒸腾流向上迁移,最终卸载至叶片中贮存[35]。
有机酸对稀土在植物中的转运起着重要的作用[64]。研究发现,水培条件下添加乙酸、苹果酸、柠檬酸能够大幅度提高大麦茎、叶中La的浓度[39],说明小分子有机酸能够通过与稀土络合并促进稀土在木质部导管中的向上运输。另有研究显示,小麦木质部伤流液中和叶片中显著富集重稀土,由于重稀土与有机配体的结合能力强于轻稀土,推测木质部汁液中有机配体的络合作用可能是导致轻、重稀土在向上迁移过程中发生分异的主要原因。Yuan等[23]发现在赣南稀土矿区生长的美洲商陆根-茎转运过程中稀土分异不明显,而茎—叶迁移过程中导致叶片重稀土富集。进一步实验发现,美洲商陆木质部汁液中柠檬酸与叶片中的稀土含量变化呈显著正相关性,证实了柠檬酸参与美洲商陆木质部中稀土的长距离运输,促进叶片中稀土的贮存[54]。此外,Shan等[40]发现添加外源组氨酸后,蕨类植物叶片中轻稀土的含量略有增加,暗示游离组氨酸可能参与轻稀土从根到叶的转运。天冬氨酸、天冬酰胺、组氨酸和谷氨酸等氨基酸也被报道可能与番茄木质部中La的长距离转运有关[65]。添加这些氨基酸均促进了番茄根对La的吸收及地上部对La的积累[65]。
除了木质部伤流液中稀土形态不同以外,转运过程造成的稀土分异还可能与植物稀土积累能力以及根—地上部的稀土分配有关。如小麦等非富集植物将更多的稀土截留在根部,细胞壁吸附和磷酸盐沉淀很大程度控制着根中有效态稀土浓度和形态[47],因此也部分决定了向上转运的稀土如何发生分异;而超富集植物如芒萁将绝大部分的稀土转运至地上部,较弱的根部区室化作用可能导致更多的轻稀土在木质部导管中随蒸腾流迁移至地上部贮存。
以上研究说明植物迁移路径中配体的种类和浓度及区室化机制是影响稀土在不同器官中分异程度的重要因素。因此,通过提取木质部伤流液,测定其中稀土形态组成及其分异特征,进而研究叶细胞中稀土的分布,将有助于揭示植物对稀土的转运机制。
4 稀土在植物地上部的亚细胞分布与解毒
稀土超富集植物与普通植物的显著不同在于稀土进入根系后大部分被转运并贮存在地上部。由于稀土比较难以进入细胞,植物叶片中的稀土大部分被吸附在质外体细胞壁上。张智勇等[66]发现芒萁叶中70%以上的稀土存在于果胶质、纤维素中,说明稀土主要分布于细胞壁中,果胶对芒萁超量积累稀土起主要作用。Shan等[40]也发现芒萁叶片中80%左右的轻稀土结合于细胞壁,其原因可能是细胞壁上带有自由羧基或羟基的多糖能够有效结合轻稀土[67]。而且在芒萁叶中细胞器、细胞质及液泡中也均有发现稀土沉积[68]。这些区室化机制使植物在较高浓度的稀土胁迫下也能正常生长,说明稀土在地上部亚细胞水平上的分布及区室化过程决定着植物对稀土的耐受程度,这也是超富集植物能够超富集稀土的原因之一。
研究发现,稀土可能与叶绿素结合,从而对植物解毒起着一定的效用。最早在蕨类植物芒萁中发现了配合物叶绿素-La和配合物叶绿素-Ce的存在[69],之后在红盖鳞毛蕨叶肉细胞中发现有6 0%~7 0%的La 与叶绿素结合分布在叶绿体中[31]。尽管叶绿素-稀土配合物在天然植物体内含量很少,但尚不能否认其存在。叶绿素与稀土形成的配合物可以部分替代叶绿素—镁的功能,甚至可能对植物的光合作用过程起促进作用[70]。此外,还有研究推测蛋白质或多肽可通过与稀土结合,使稀土失活或者对植物细胞代谢的毒性降低,参与稀土的解毒和稳态,在稀土超富集过程中发挥重要作用[71]。
5 稀土在超富集植物中分异的概念模型
综上所述,我们以芒萁和美洲商陆为代表,初步建立了超富集植物中稀土分异的概念模型(图1),主要由四个关键过程组成:
1)根际土壤溶液化学过程:植物根系主要吸收自由离子态稀土,根际有机配体的种类和浓度、pH、有机质和离子扩散等因素,会通过络合、吸附、解吸和沉淀等作用降低或者提高了土壤溶液中稀土生物有效性,影响根系对稀土的吸收,进而使稀土在植物吸收过程中产生分异。
2)根系吸收:稀土通常以自由离子态形式吸收进入植物根系。芒萁和美洲商陆根系可能通过Ca2+通道等低亲和力运输系统被动吸收稀土,其中美洲商陆根系还能通过Al转运蛋白等高亲和力运输途径吸收稀土。这些吸收途径可能使芒萁根系富集轻稀土,美洲商陆根系轻微富集轻稀土。
3)木质部转运:稀土随着木质部流动向上转运。其中芒萁较弱的根部区室化作用可能导致更多的轻稀土在木质部导管中以自由离子态形式随蒸腾流迁移至地上贮存。相反,美洲商陆木质部中重稀土更易与有机酸等络合并向地上部转运,尤其柠檬酸在木质部重稀土的长距离转运过程中起着重要作用。
4)地上部分布解毒:最终大量稀土贮存在植物叶片中。芒萁地上部中,稀土在叶柄向叶片转运时轻稀土与重稀土的比值最大,叶片中轻稀土大部分被细胞壁吸收保持在质外体中,或被卸载至液泡中,也可能进入细胞与蛋白质、叶绿素等络合进行解毒。美洲商陆叶片中解毒情况目前尚不清楚,推测可能存在与芒萁类似的区室化机制进行解毒,使重稀土大量富集在叶片中。

图1 稀土在超富集植物中的积累、分布与分异示意图Fig. 1 Schematic diagram of accumulation, distribution and fractionation of REEs in hyperaccumulators
6 展 望
稀土元素在土壤-植物体系中迁移与分异的理论体系仍然不健全,因此需要从下面几个方面展开更深入的研究:
1)深入了解影响超富集植物根际稀土生物有效性的关键控制过程。如超富集植物根系是否分泌影响根际稀土生物有效性的小分子有机酸,这些小分子有机酸如何影响根际稀土的形态和分异特征,从而导致稀土向植物迁移过程中产生分异。根表是否存在由扩散介导的稀土络合物解离过程,使根表附近自由离子态稀土浓度发生变化,从而影响根系对稀土的吸收。
2)根部质外体途径运输不利于稀土向地上部转运,因此稀土超富集植物理应存在高效的共质体运输途径,然而到底存在哪些跨膜运输通道以及方式尚不清楚,可以结合根际稀土形态和根系稀土分异特征,进一步开展根系吸收动力学、分子蛋白机制等研究,从关键基因的角度揭示超富集植物根系对稀土的高效吸收策略。
3)进一步研究稀土在木质部中以什么形态被高效转运至地上部,生物大分子和有机小分子在稀土转运过程中起什么作用,是否可以通过木质部伤流液中稀土形态组成及其分异特征去模拟这一过程?
4)稀土在叶片亚细胞水平的分布与解毒如何?除了以磷酸盐稀土沉淀形式存在于细胞壁中进行解毒,是否存在稀土与小分子有机酸结合产生无毒或低毒复合物区室化于叶片上下表皮细胞和液泡中进行解毒?
5)进一步探索稀土超富集植物的分异模式对稀土吸收和转运机理的指示作用。
