基于表型性状和CDDP分子标记延安万花山牡丹遗传多样性分析
2019-08-13翟立娟史倩倩李想
翟立娟 史倩倩 李想

摘要:利用28个表型性状和CDDP分子标记对延安万花山牡丹进行遗传多样性分析。结果表明,万花山牡丹的表型性状存在较大差异。质量性状的香农多样性指数变化范围为0.28~1.21,其中花色、叶端形态、叶片颜色、花型等较高;9个数量性状在样本中差异极显著,变异系数变化范围为12.10%~69.23%,其中花瓣数、雄蕊数的变异系数较大。CDDP分子标记分析结果表明,万花山牡丹有效等位基因数为1.599 4,Neis基因多样性指数和香农多样性指数分别为0.333 5和0.486 1,表明延安万花山牡丹具有较高的遗传多样性。2种分析方法结果具有一致性,均可将延安万花山牡丹分为三大类,根据与紫斑牡丹和矮牡丹亲缘关系可划分为3个类型,即接近矮牡丹的类型、接近紫斑牡丹的类型和中间类型。
关键词:牡丹;遗传多样性;表型性状;CDDP分子标记
中图分类号: S685.110.24 文献标志码: A 文章编号:1002-1302(2019)02-0095-06
延安万花山位于陕西省延安市城区西南方向,长期栽培多种牡丹品种,这些牡丹构成中原牡丹品种群延安牡丹亚群[1-2]。延安万花山有延安牡丹(Paeonia yananensis)、矮牡丹(P. jishanensis)及紫斑牡丹(P. rockii)的野生居群[3-5]。关于延安万花山牡丹类群的起源及物種地位有多种看法,郑宏春等将万花山上的牡丹划分为4个变型,即白花矮牡丹、紫斑矮牡丹、红花矮牡丹和紫斑红花矮牡丹[6]。洪涛等将其中小叶数11枚、花粉色、花瓣有紫黑色瓣块的牡丹提升到种的级别,即延安牡丹[3]。洪德元等认为,延安牡丹是矮牡丹和紫斑牡丹的杂交后代[7],后人利用RAPD分子标记[8]、孢粉学[9]、形态学标记[4]、cpDNA序列和SSR分子标记[10]得到类似的结论。此地牡丹长期处于半野生半栽培状态,在花型、花色等性状方面存在较大变异[11],是挖掘观赏植物重要性状的关键基因和牡丹新品种培育的良好试材[12]。李嘉珏的《中国牡丹》记载,该地至少有16个品种。然而,笔者调查发现该地牡丹品种数量正急剧减少,某些品种仅剩几株,甚至不见踪迹,如不采取保护措施,这将是牡丹育种栽培资源的一大损失,同时发现万花山上保存着一些仍未命名或品种名遗失的牡丹样本。目前缺乏关于延安万花山牡丹品种及其遗传多样性的研究,这不利于该地牡丹资源的保存和合理开发利用。
表型多样性是了解遗传变异的重要线索[13],具有操作简单、直观、便于观察等优点[14-15]。在牡丹野生种[16-17]和栽培品种[18]的遗传多样性研究中,得到了较可靠的结果。保守DNA序列多态性(conserved DNA-derived polymorphism,CDDP)分子标记是一种基于DNA保守序列的新型分子标记方法,具有稳定性、重复性好的优势,能产生丰富的遗传信息[19],已在水稻[20]、菊花[21]、栽培牡丹[22]等植物遗传多样性研究中得到广泛应用。因此,这2种方法的结合,可以从表型和基因型2个维度去准确、细致地了解其遗传多样性。
本试验以分布于延安万花山的24个牡丹样本(包括10个形态能够稳定遗传但尚未命名或品种名遗失的样本、3个当地自然野生状态下的牡丹样本和11个已被记载的牡丹品种)为研究对象,以延安市富县分布的紫斑牡丹和山西省稷山县的矮牡丹为对照,采用28个表型性状分析和CDDP分子标记技术相结合的方法研究延安万花山牡丹的遗传多样性,以期为该地牡丹种质资源的保护和合理开发利用提供理论依据。
1 材料与方法
1.1 试验材料
2014—2016年连续3年于牡丹花期对延安万花山牡丹进行资源调查,对性状能够稳定遗传且开花稳定的24个类型进行表型性状记录,取每个类型的3~5个植株嫩叶用于DNA提取;同时以延安市富县张家林场的紫斑牡丹(108.76° E、36.36° N,海拔1 461 m)和山西省稷山县的矮牡丹(35.72° N、110.95° E)为对照进行试验。试验样品的相关信息见表1和图1,其中1~10号样本尚未命名或品种名遗失,11~13号为万花山当地自然野生状态下的牡丹样本,14~24号为已被记载的牡丹样本[1,11],25~26号为富县紫斑牡丹,27号稷山县的矮牡丹。
1.2 试验方法
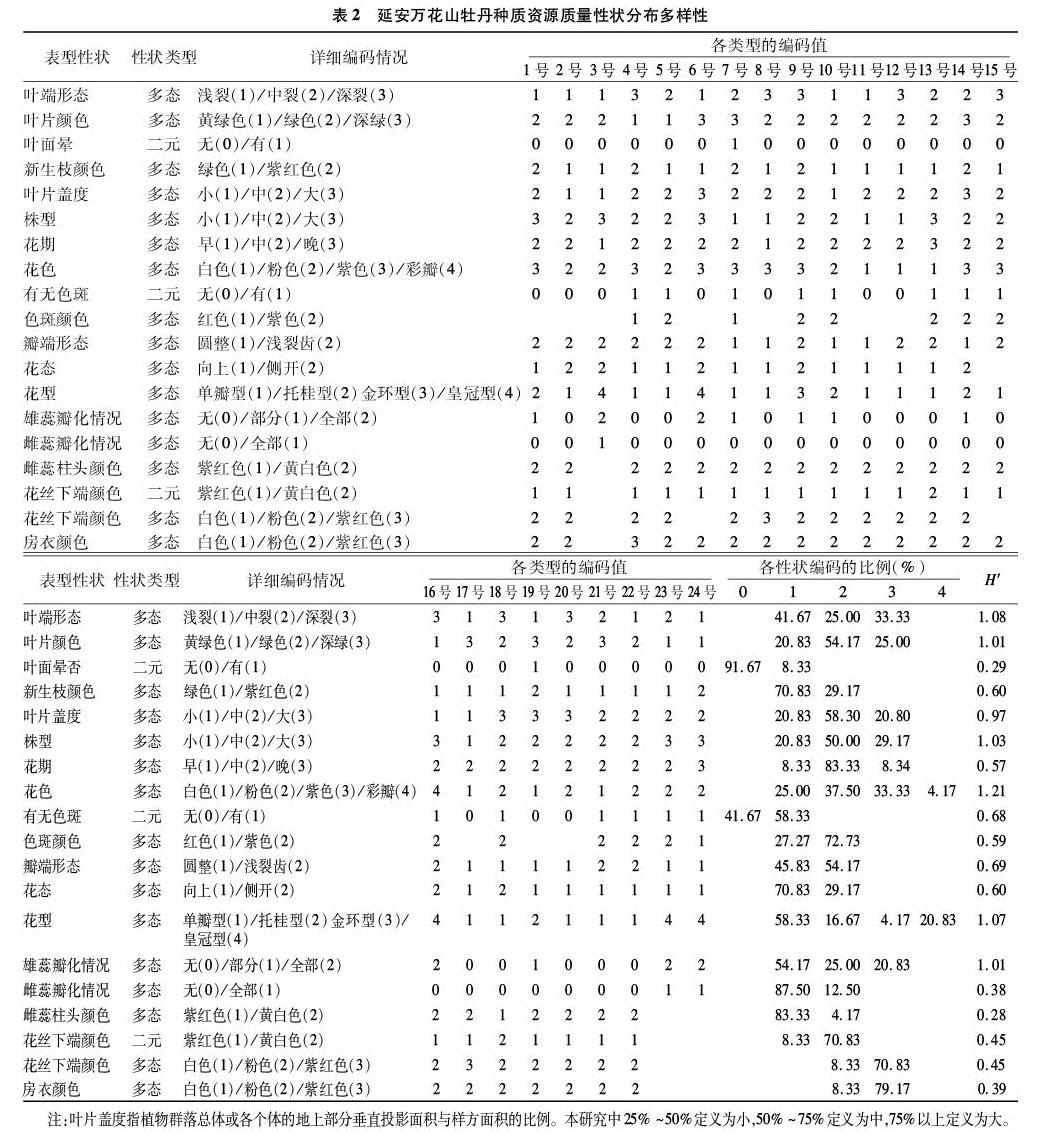
参照成仿云等的方法[2,15,18]选择具有代表性、遗传相对稳定的28个表型性状,其中19个质量性状及赋值标准见表2,9个数量性状见表3。利用改良的CTAB法[23]提取27份材料的DNA,然后参照李莹莹方法[19]利用14条CDDP引物进行PCR扩增,PCR产物在2%琼脂糖凝胶上电泳分离,凝胶成像系统观察和保存电泳图谱。
1.3 数据处理与统计分析
表型性状数据统计中,利用SPSS 22.0软件进行质量性状的分布频率分析、数量性状单因素方差分析、采用Euclidean距离、Ward聚类方法进行Q型聚类[18]。各性状的遗传多样性采用香农多样性指数(H′)进行评价。对于CDDP带型,首先按照电泳图谱中同一位置上DNA条带的有无进行统计,形成0/1矩阵,然后应用POPGEN 32计算供试材料的有效等位基因数、Neis基因多样性指数、香农多样性指数,最后运用NTSYS软件进行UPGMA法构建聚类树状图,其余计算由Excel 2007完成。
2 结果与分析
2.1 表型性状遗传多样性分析
24个延安万花山牡丹样本质量性状分布及多样性情况见表2,香农多样性指数(H′)的变化范围为0.28~1.21,平均值为0.70。其中,多样性指数最高的质量性状是花色(1.21),主要以白色(25%)、粉色(37.5%)、紫红色(33.33%)及复色(4.17%)为主;共有6个质量性状的多样性指数大于1, 包括叶端形态(1.08)、叶片颜色(1.01)、株型(1.03)、花型(1.07)及雄蕊瓣化情况(1.01);叶面晕(0.29)、雌蕊瓣化(0.38)及房衣颜色(0.39)的香农多样性指数(H′)相对较低;柱头颜色的遗传多样性指数最低(0.28)。
F值说明了分析结果可靠程度和不同性状在不同样品间的差异程度。如表3所示,在群体水平上,延安万花山牡丹的9个数量性状的差异均达到极显著水平(P<0.01),表明9个性状在群体间层次上存在广泛的变异。花瓣数在群体间F值最大(94.50);雄蕊数和花径的群体间F值较大,分别为81.90和45.98;外花瓣数的群体间F值最小(7.53)。多重比较结果(表3)显示,尽管各数量性状在居群间差异明显,但是大多表现为连续变异。
数量性状变异系数(CV)表示性状的离散程度,反映表型的变异特征。延安万花山牡丹种质资源的数量性状(表3)变异系数变化范围为12.10%~69.23%。其中花瓣数的变异系数最大,为69.23%,雄蕊数的变异系数较大,大于60%;小叶数量的变异系数(27.17%)、复叶长的变异系数(16.70%)、花径的变异系数(13.31%)和着花量的变异系数(22.74%)均较小;叶柄长的变异系数最小,为12.10%。
根据28个表型性状对所有材料进行Ward聚类分析,结果见图2,在遗传距离为15.5时,供试材料被分为3个大类,第Ⅰ大类包含2份紫斑牡丹,它们小叶数量在30枚以上,花丝为白色,心皮为黄白色;第Ⅱ大类包含22份样本,包括了延安万花山大部分牡丹样本和矮牡丹,当地的矮牡丹(11、12号样本)与稷山县矮牡丹的亲缘关系较近;第Ⅲ大类包含3份样品,它们均为粉色,花型为皇冠型。在遗传距离为9.5时,第Ⅱ大类的样本又可以分为4类,第ⅰ类包含万花山7号和万花春(19号)2份样本,它们的雄蕊均有少量瓣化;第ⅱ类包含6份万花山牡丹样本和1个矮牡丹,其均为单瓣型,花瓣基部没有色斑;第ⅲ类包含7个样本,其花瓣基部均有色斑,万花山5号、瑞香紫(15号)、万花山4号聚在一起,它们均为紫色花且花型为单瓣型;第ⅳ类包含6个样本,除粉盘黄珠(18号)花型为单瓣型外,其余样本花型为重瓣型。总体上讲,大部分延安万花山牡丹样本在形态上更接近于矮牡丹。
2.2 CDDP的遗传多样性分析
利用14条CDDP引物对27份牡丹材料进行扩增,每条引物扩增出的条带数、多态性条带数和比例及其特异性条带和比例均有较大的差异(表4),表明CDDP分子标记的多态性较高。利用POPGEN 32分析延安万花山牡丹遗传多样性,得到有效等位基因数为1.599 4,Neis基因多样性指数为 0.333 5,Shannon-Wiener信息指數为0.486 1,多态性位点数为109,多态性位点率为85.2%。结果表明延安万花山的牡丹多态性较高。
根据Neis遗传一致度,利用UPGMA法进行聚类分析(图3)。在遗传距离0.718处,可将样本分为3个类群。其中,第Ⅰ类群包括14个万花山牡丹和矮牡丹,其中的矮牡丹与万花山11号的亲缘关系最近。第Ⅱ类群包括9个样本,2个野生紫斑牡丹属于这一类群,此类群中样本花瓣基多有色斑;第Ⅲ类群包括4个样本,3号和10号亲缘关系最近,之后与12号和5号样本聚在一起。
2.3 表型性状和CDDP分子标记的相关分析
在本研究中,2种方法的分析结果虽有差异,但也有一定的相关性。2种方法的香农多样性指数(H′)分别为0.70和 0.486 1,说明延安万花山牡丹具有较高的遗传多样性。2种方法均可把27份样本分为三大类,花瓣基部没有色斑的多与矮牡丹聚在一起,花瓣基部有色斑的多于紫斑牡丹聚在一起;同一色系或相近花型的延安万花山牡丹样本遗传距离较近。在2种方法的聚类树状图中,万花山9号和延州红、狮子头和延州粉均聚在一起,说明它们彼此之间亲缘关系比较近。
3 讨论与结论
研究牡丹遗传多样性的方法有形态学标记、细胞学标记、生理生化标记和分子标记[24],其中形态学标记和分子标记使用最广泛[25-27]。本研究中,延安万花山牡丹种质资源的28个表型性状存在明显差异。CDDP分子标记获得的Neis基因多样性指数,香农多样性指数多态性位点的比例均较高。2种方法均说明延安万花山牡丹具有较高的遗传多样性。但2种方法的聚类结果也存在一定的差异,一方面是因为表型性状是样品的基因型值、环境效应、基因型与环境互作效应的综合结果,而分子标记检测的多态性只是DNA水平的变异;另一方面可能与其杂交起源遗传背景有关。因此2种方法结合,才能更加准确地分析种群的遗传变异状况。
延安牡丹是临近区域分布的紫斑牡丹和当地矮牡丹的杂交后代[10,12]。根据形态学分析,矮牡丹植株较矮,小叶多为9枚,近圆形或卵形,因顶生小叶分裂程度不同而表现9、11、15枚的变化;紫斑牡丹植株较高大,小叶数15枚及以上,多呈卵状披针形,花色为白色、粉色和紫红色[3-5];延安牡丹同时具有2个亲本的性状,小叶数11(13)枚,小型圆叶,花瓣基部具有紫黑色斑纹,主模式为瑞香紫中的1株[3,11]。本研究发现万花山还存在许多类似于延安牡丹的半野生半栽培的牡丹样本,小叶数15枚以上并呈卵状(万花山11号)、花瓣高度瓣化(万花山3号)、花瓣基部有红斑并且叶面有红晕(万花山7号)的类型或品种,明显不同于矮牡丹和紫斑牡丹的性状,说明延安万花山牡丹存在丰富的变异,可以推断为紫斑牡丹与矮牡丹的杂交后代只是在形态上出现了连续变异[28]。根据聚类结果,可以把万花山的牡丹分为接近矮牡丹的类型,接近紫斑牡丹的类型及中间类型。接近矮牡丹的类型大致包括2、7、8、11、12、17、19、20号材料,它们多为单瓣型,花瓣基部没有色斑;接近紫斑牡丹的类型包括4、13、14、15、18、21、22
号材料,它们花瓣基部多有色斑;中间类型包括1、3、5、6、9、10、16、23、24号材料,这些材料多为重瓣花型。对于延安万花山牡丹的杂交机制还有待进一步研究。
本研究结果表明延安万花山牡丹存在丰富的变异,具有较高的观赏和科研价值。重瓣类型、具有色斑的类型、雄蕊少量瓣化的类型均可作为挖掘观赏植物重要性状的关键基因的重要种质资源。然而,目前延安万花山牡丹种质资源流失严重,应该对万花山地区的牡丹进行全面的资源清查和品种登记,加大科研力度,积极利用最新的育种和杂交技术,培育以延安万花山地区的牡丹为基础的新品种。
参考文献:
[1]李嘉珏. 中国牡丹与芍药[M]. 北京:中国林业出版社,1999:80-81.
[2]成仿云,李嘉珏. 中国紫斑牡丹[M]. 北京:中国林业出版社,2005:87-94.
[3]洪 涛,张家勋,李嘉珏,等. 中国野生牡丹研究(一)芍药属牡丹组新分类群[J]. 植物研究,1992,12(3):223-234.
[4]袁 涛,王莲英. 我国芍药属牡丹组革质花盘亚组的形态学研究[J]. 园艺学报,2003,30(2):187-191.
[5]洪德元. 紫斑牡丹及其一新亚种[J]. 植物分类学报,1998,36(6):538.
[6]郑宏春,陈 凯,李明瑞. 万花山矮牡丹及其变型的初步研究[J]. 西北植物学报,1992,12(7):142-146.
[7]洪德元,潘开玉. 芍药属牡丹组的分类历史和分类处理[J]. 植物分类学报,1999,37(4):351.
[8]邹喻苹,蔡美琳,王子平. 芍药属牡丹组的系统学研究——基于RAPD分析[J]. 植物分类学报,1999,37(3):220-227.
[9]袁 涛,王莲英. 几个牡丹野生种的花粉形态及其演化、分类的探讨[J]. 北京林业大学学报,1999,21(1):17-21.
[10]Yuan J H,Cheng F Y,Zhou S L. Hybrid origin of Paeonia×yananensis revealed by microsatellite markers,chloroplast gene sequences,and morphological characteristics[J]. International Journal of Plant Sciences,2010,171(4):409-420.
[11]李嘉珏,张西方,赵孝庆. 中国牡丹[M]. 北京:中国大百科全书出版社,2011:300-303.
[12]袁军辉. 紫斑牡丹及延安牡丹起源研究[D]. 北京:北京林业大学,2010.
[13]杨生超,徐绍忠,文国松,等. 灯盏花种质资源群体表型多样性研究[J]. 西北植物学报,2008,28(8):1573-1579.
[14]刘 胤,陈 涛,张 静,等. 中国樱桃地方种质资源表型性状遗传多样性分析[J]. 园艺学报,2016,43(11):2119-2132.
[15]张晓骁,宋 超,張延龙,等. 秦岭与子午岭地区紫斑牡丹居群表型多样性研究[J]. 园艺学报,2017,44(1):139-150.
[16]李树发,蔡艳飞,张秀新,等. 滇牡丹天然群体的表型多样性[J]. 西南农业学报,2016,29(10):2470-2478.
[17]李林昊,张延龙,牛立新,等. 秦岭地区‘凤丹牡丹居群果期相关性状的表型多样性研究[J]. 西北林学院学报,2015,30(4):127-131.
[18]韩雪源,张延龙,牛立新. 9个牡丹品种的形态学分类研究[J]. 西北农林科技大学学报(自然科学版),2014,342(9):128-136.
[19]李莹莹,郑成淑. 利用CDDP标记的菏泽牡丹品种资源的遗传多样性[J]. 中国农业科学,2013,46(13):2739-2750.
[20]Collard B,Mackill D J. Conserved DNA-Derived polymorphism (CDDP):a simple and novel method for generating DNA markers in plants[J]. Plant Molecular Biology Reporter,2009,27(4):558-562.
[21]李 田,郭俊娥,郑成淑,等. 菊花品种的遗传多样性分析及CDDP指纹图谱构建[J]. 北京林业大学学报,2014,36(4):94-101.
[22]Wang X W,Fan H M,Li Y Y,et al. Analysis of genetic relationships in tree peony of different colors using conserved DNA-derived polymorphism markers[J]. Scientia Horticulturae,2014,175(1):68-73.
[23]何雪娇,郑 涛,苏金强,等. 改良CTAB法提取野牡丹科7种植物DNA[J]. 广东农业科学,2011,38(18):120-122.
[24]Ji L J,Wang Q,da Silva J A,et al. The genetic diversity of Paeonia L[J]. Scientia Horticulturae,2012,143(1):62-74.
[25]Budak H,Shearman R C,Parmaksiz I,et al. Molecular characterization of buffalograss germplasm using sequence-related amplified polymorphism markers[J]. Theoretical and Applied Genetics,2004,108(2):328-334.
[26]Duan Y B,Guo D L,Guo L L,et al. Genetic diversity analysis of tree peony germplasm using iPBS markers[J]. Genetics&Molecular Research Gmr,2015,14(3):7556-7566.
[27]Homolka A,Berenyi M,Burg K,et al. Microsatellite markers in the tree peony,Paeonia suffruticosa(Paeoniaceae)[J]. American Journal of Botany,2010,97(6):E42-E44.
[28]郝津藜,赵 娜,石颜通,等. 黄牡丹远缘杂交亲和性及杂交后代形态分析[J]. 园艺学报,2014,41(8):1651-1662.
