竹节参转录组使用密码子偏好性分析
2019-08-13梁娥齐敏杰丁延庆
梁娥 齐敏杰 丁延庆
摘要:竹节参是我国珍稀濒危中药材,研究其基因密码子使用模式,可为利用基因工程技术实现人参皂苷异源生物合成及竹节参分子育种改良提供理论依据。以竹节参转录组测序结果为数据来源,筛选编码蛋白基因序列(coding sequence,简称CDS)碱基数不小于300 bp的 11 199 条完整开放阅读框序列作为研究对象,利用Codon和SPSS软件分别统计竹节参基因密码子GC含量、密码子第3位的(C+G)含量(GC3)和密码子第1、第2位(G+C)含量的平均值(GC12)、同义密码子的相对使用度(RSCU)、有效密码子数(ENC)等密码子偏好性指标,通过中性绘图(GC12 vs. GC3)、PR2绘图和ENC-GC3s绘图分析影响竹节参密码子使用模式的因素。结果表明,竹节参基因的平均GC、GC12和GC3s含量分别为44.67%、46.97%和39.80%,其密码子使用模式受到突变和选择等多重因素的影响,确定了31个竹节参最优密码子,除了UUG外,其余最优密码子均以A或T结尾。竹节参密码子使用模式与大肠杆菌和酿酒酵母相比差异较大,选取毕赤酵母作为竹节参基因的异源表达宿主更为合适。
關键词:竹节参;转录组;密码子使用模式;最优密码子
中图分类号: Q755;S567.5+10.1 文献标志码: A 文章编号:1002-1302(2019)02-0059-05
遗传密码子是生物体DNA与蛋白质之间信息传递的基本单位,具有简并性,即同一氨基酸有多个对应的密码子,编码同一种氨基酸的密码子叫作同义密码子,同义密码子在同一物种不同基因间或不同物种内的使用频率大有不同,这种不均衡使用模式称为密码子使用偏好性,通常把使用频率较高的一种或几种同义密码子称为最优密码子[1-2]。研究显示,不同物种之间基因密码子偏好性是由突变压力(如GC含量、基因碱基组成)和自然选择作用(如翻译起始信号、基因表达水平、蛋白结构与长度、tRNA丰度等)引起的,mRNA的二级结构及其稳定性、翻译的速度和准确度、蛋白质折叠等因素也与密码子的偏好性有关[3-4]。对物种密码子偏好性开展研究,有助于理解物种进化发展及密码子使用偏好性的调控机制。密码子偏好性在基因异源表达研究方面也显示了重要作用,基因的表达量越大,其密码子偏好性越强[5],根据这一原理,替换基因低表达密码子可以提高外源基因表达量,同时根据密码子使用偏好性可以选择更为合适的宿主表达系统。有报道表明,可通过优化密码子的方式来提高外源基因在宿主细胞中的表达量[6-7]。周宗梁等通过优化密码子的方法提高了苏云金芽孢杆菌基因cry1Ah在玉米和水稻中的表达量[8-9];杨金玲等通过优化蝎毒镇痛活性肽基因BmKAngM1,将其导入毕赤酵母后该基因表达量得到显著提高[10];通过使用最优密码子,在草菇[11]、拟南芥[12]、川母贝[13]、菠萝[14]等生物中均得到了很好的研究成果。
竹节参(Panax japonicus C. A. Mey)为多年生草本植物,属于五加科(Paeoniaceae)人参属,是我国珍稀濒危的“七类中草药”之一,具有抗炎、延缓衰老、降血糖等药理作用,有着极高的药用和保健价值。竹节参中富含活性物质三萜皂苷,也是其特征性成分。目前在竹节参种质资源[15-16]、毛状根的培养[17]、基因工程代谢的调控[18]、生药学鉴定[19]、精油成分分析[20]和三萜皂苷代谢追踪[21]等几个研究领域已经开展了许多研究工作,但直接从竹节参中通过分离提取的方法获得三萜皂苷对资源消耗极大,技术难度较高。若利用生物合成的方法则可以很好地解决这一困难,实现有效成分的生物合成,选择适合关键酶基因高效表达的异源表达系统是一个重要步骤。本研究以竹节参转录组数据为材料,通过分析竹节参基因密码子组成的各项指标,研究竹节参表达基因密码子使用偏好性及其影响因素,以期为竹节参相关基因表达系统的选择及分子育种提供理论基础。
1 材料与方法
1.1 数据来源
竹节参转录组数据来源于文献[22],通过Perl语言程序对竹节参转录组数据进行过滤筛选,筛选出碱基数≥300 bp的蛋白质编码序列共11 199条,作为密码子分析的数据来源。本研究中使用到的大肠杆菌(Escherichia coli)、酿酒酵母(Saccharomyces cerevisiae)和毕赤酵母(Pichia pastoris)的密码子偏好性数据来自Codon Usage database( http://www. kazusa. or. jp /codon /)。
1.2 竹节参基因GC含量分析及中性绘图
利用Codon W1.4.2(http://codonw.sourceforge.net/)统计分析竹节参基因密码子的碱基组成规律,测得鸟嘌呤和胞嘧啶总体含量G+C、密码子第3位碱基组成(A3、G3、C3、T3)、密码子第3位的C+G数(GC3)和密码子第1、第2位的平均G+C含量(GC12)等数据。本研究以密码子第1、第2个碱基的G+C平均含量为纵坐标,以密码子第3个碱基的G+C含量为横坐标,通过中性绘图探究影响密码子偏好性的因素。分析密码子第1、第2位与第3位碱基组成的相关性,来分析自然选择与压力突变对基因密码子使用模式的影响[23]。
1.3 竹节参基因有效密码子数分析
参照Fuglsang的方法[24],利用有效密码子数来衡量竹节参基因单个密码子使用的偏好程度。基因有效密码子数(effective number codon,简称ENC)是衡量基因密码子偏好性的一个重要指标,数值范围为20(每个氨基酸只使用1个同义密码子的极端偏好情况)到61(每个同义密码子被平均使用的无偏好情况)。研究表明,当ENC值≤35时,基因密码子的使用偏好性越强[25]。同时以ENC值为纵坐标,GC3s为横坐标进行ENC绘图,分析各基因密码子的使用特征,研究竹节参密码子偏好性和基因碱基组成之间的关系。
1.4 同义密码子相对使用度及最优密码子分析
参照Sharp等的方法[26],采用同义密码子的相对使用度(RSCU)作为衡量竹节参密码子使用偏好性的指标。使用3个指标表示:密码子使用无偏好性(RSCU=1);密码子使用度较大(RSCU>1);密码子使用频率较低(RSCU<1)。利用高表达优越密码子分析方法[27],统计所有基因的ENC值、有序数据集的上下10%区间形成高RSCU集合和低RSCU集合,进行最优密码子分析。根据2个子集的ΔRSCU值及卡方检验确定最优密码子。
1.5 PR2绘图分析
采用PR2绘图进行密码子奇偶偏好分析[23],分别统计竹节参基因中A3/(A3+T3)与G3/(G3+C3)数据,以此为纵坐标和横坐标,通过平面图显示各基因碱基的组成。为使编码密码子第3位碱基对A/T或T/A和G/C或C/G的突变不均衡,PR2绘图时只选择亮氨酸(Leu,密码子:CTA、CTC、CTG、CTT)、脯氨酸(Pro)、精氨酸(Arg,密码子:CGA、CGC、CGG、CGT)、丝氨酸(Ser,密码子:TCA、TCC、TCG、TCT)、丙氨酸(Ala)、缬氨酸(Val)、苏氨酸(Thr)和甘氨酸(Gly)等遗传密码子进行分析。
2 结果与分析
2.1 GC含量分析以及中性绘图
竹节参转录组数据中共有11 199条完整的开放阅读框序列,利用Codon W对完整的开放阅读框序列进行密码子使用模式分析。结果(表1)表明,所有完整的开放阅读框序列总长度为 12 006 732 bp,N50=1 332 bp,平均GC含量为44.67%,GC含量范围为30.3%~63.8%。竹节参基因组平均GC含量低于大肠杆菌基因组平均GC含量(52.35%),高于酿酒酵母基因组平均GC含量(39.77%)和毕赤酵母基因组平均GC含量(42.73%)。竹节参基因组密码子第1、第2位碱基GC含量的变化范围为31.7%~71.8%,平均GC含量为46.97%。密码子的第3位上碱基GC含量的变化范围为10.7%~83.9%,平均GC含量是39.80%。密码子第3位碱基上A、T的使用频率分别是33.09%、41.27%,略高于C、G的使用频率(分别为23.61%、27.57%),说明竹节参基因密码子对以A和T结尾的密码子的使用偏好程度较大,而对以G和C结尾的密码子的使用偏好程度较小。密码子第3位碱基上的平均GC含量(39.80%)略低于毕赤酵母基因密码子第3位碱基的平均GC含量(42.16%),偏高于酿酒酵母密码子第3位碱基的平均GC含量(38.10%),大大低于大肠杆菌遗传密码子第3位碱基的GC平均含量(55.62%)。研究表明,竹节参对密码子使用并无特殊碱基偏好,其密码子使用模式与大肠杆菌和酿酒酵母差异较大,与毕赤酵母差异略小。如图1所示,中性绘图分析表明,竹节参密码子GC12取值范围为31.7%~71.8%,GC3的取值范围为10.7%~83.9%,GC3s与GC12的相关系数为0.062,回归系数为0.48,表明并无显著相关性,说明竹节参基因密码子使用模式主要受到自然选择的影响。
2.2 竹节参基因有效密码子数分析
研究显示,竹节参ENC值一般在25.88~61.00,ENC平均值为53.04。通常将ENC值=35作为衡量密码子偏好性强弱的标准[22]。竹节参基因有13条ENC值小于35,表明竹节参基因整体水平密码子偏好性较低,只有少数基因具有密码子偏好性。由表2的ENC相关性分析可知,GC与GC12、GC3之间的相关性均达极显著水平,GC12和GC3之间并没有明显的相关性,密码子成分不相同,而ENC值与密码子数的相关性也没有达到显著水平,说明密码子数对ENC的影响很弱,排除了基因长度过短对密码子偏好性的影响。如图2所示,ENC-GC3s绘图分析结果表明大部分竹节参基因均分布在标准曲线的周围,而小部分基因则分布在距标准曲线较远的位置,说明竹节参表达基因密码子使用模式不仅受到自然选择的影响,同时也受到突变压力作用的影响。
2.3 同义密码子的相对使用度及最优密码子分析
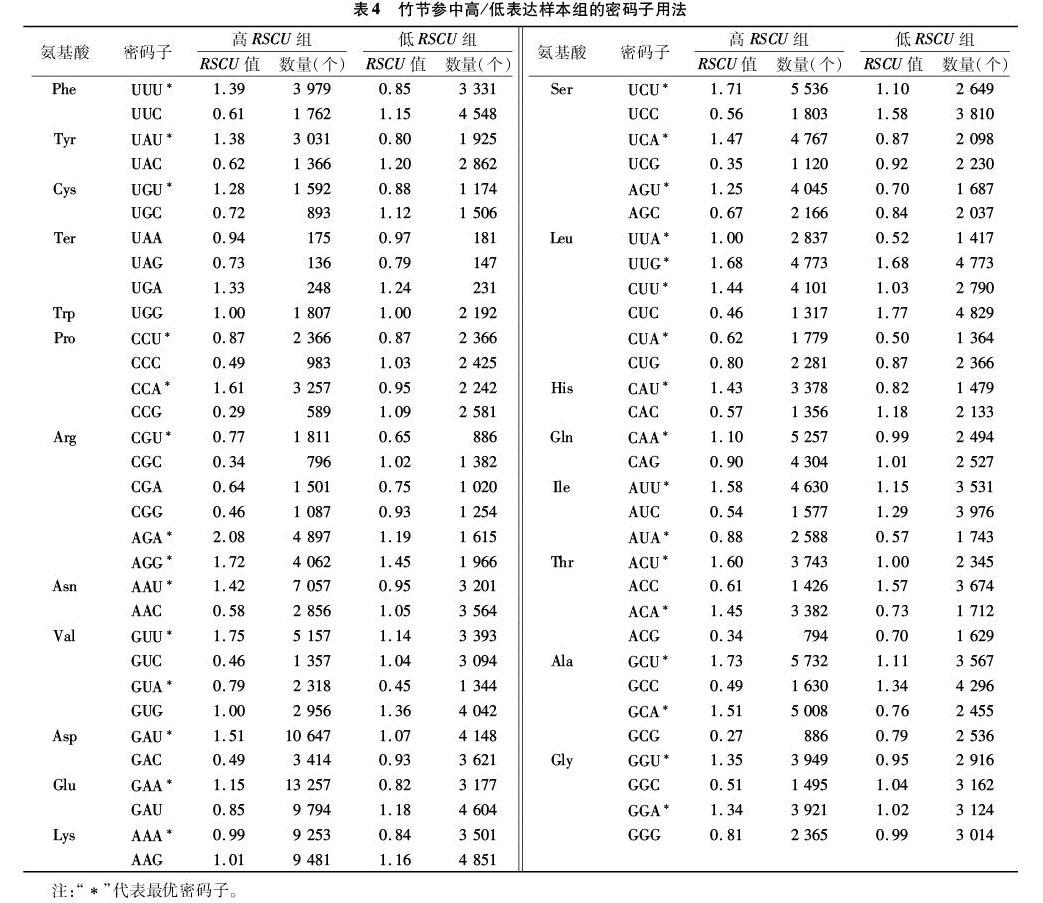
如表3所示,竹节参共有28个密码子的RSCU值大于1,说明这28个密码子可能是竹节参偏好使用的。除去Trp和Met(只有1种密码子编码)外,编码Phe、Ser、Leu、Tyr、Cys、His等氨基酸的密码子及终止密码子存在较大的偏好性,RSCU>1的密码子主要以U和A结尾。如表4所示,对竹节参基因进行高、低表达样本库比较,筛选出竹节参转录组样本的最优密码子共31个,分别是UUU、UUA、UUG、CUU、CUA、AUU、AUA、GUU、GUA、UAU、CAU、CAA、AAU、AAA、GAU、GAA、UCU、UCA、CCU、CCA、ACU、ACA、GCU、GCA、UGU、CGU、AGU、AGA、AGG、GGU、GGA,它们分别编码苯丙氨酸、亮氨酸、异亮氨酸、缬氨酸、酪氨酸、组氨酸、谷氨酰胺、天冬酰胺、赖氨酸、天冬氨酸、谷氨酸、丝氨酸、脯氨酸、苏氨酸、丙氨酸、半胱氨酸、精氨酸和甘氨酸共18个氨基酸,其中Arg有4个最优密码子。在这31个最优密码子中,除了UUG是以G结尾外,其他密码子均是以A或T结尾的,说明竹节参偏好使用以A/T结尾的密码子。
2.4 PR2-plot分析
通过PR2-plot分析,表明了竹节参各基因密码子中4个碱基组分嘌呤(A和G)与嘧啶(T和C)之间的关系,如图3所示,4个碱基均不均衡使用,竹节参密码子第3位碱基T的使用频率大于碱基A,碱基G的使用频率大于碱基C,表明竹节参基因密码子使用模式受到突变压力和自然选择等多重因素的影响。
3 讨论
每种生物的特有密码子使用模式是物种为适应自然环境而进化形成的。作为生物基因组中碱基组成的一项重要指标,GC含量的高低表明了生物突变方向性的频数大小。研究表明,密码子第3位上GC碱基的含量受到突变压力的影响较小,因此GC3s含量通常被作为分析密码子使用模式的一个主要参数。本研究分析竹节参基因GC含量分布情况,发现平均GC含量和GC12比较接近(均低于50%),表明所有竹节参基因中整体AU含量略高于GC,且密码子主要以A和U结尾。相比于毕赤酵母,其密码子使用模式与大肠杆菌和酿酒酵母的差异更大。多数研究者对高等植物如水稻(Oryza sativa)、小麦(Triticum aestivum)、大麦(Hordeum vulgare)、真菌与一些单细胞微生物如镰状疟原虫(Plasmodium falciparum)、支原体(Mycoplasmacapricolum)和植物线粒体微基因组中密码子的使用模式进行了研究,结果表明高等植物基因密码子中GC含量相对较高且趋于以G/C结尾[25],在低等生物中则AU含量明显高于GC含量[26-31]。研究表明,物种密碼子使用模式在形成过程中往往受到很多因素的影响,如突变压力和自然选择等。本研究中中性绘图分析结果表明,GC12与GC3s二者之间其实不存在明显的相关性;ENC-plot分析表明,竹节参基因大部分均分布在标准曲线周围,而小部分基因则分布于距标准曲线较远的位置;PR2-plot 分析发现,竹节密码子A、C、T和G 4个碱基使用不均衡,结合中性绘图、ENC-plot和 PR2-plot综合分析表明,竹节参基因密码子使用模式受到突变压力和自然选择等多重因素的影响。通过高偏性与低偏性基因库RSCU值的比较,利用ΔRSCU大小等级划分以确定最优密码子[32-35],本研究中通过此方法筛选出竹节参基因偏好密码子共31个,在31个密码子中,除了UUG外,其余最优密码子均以A或T结尾,以此推测竹节参基因密码子的偏好性与GC3s含量呈负相关。有研究者对长春花[36]、小麦[23]等植物密码子使用模式进行研究发现,最优密码子大多是以A/T结尾的,且通常表现出对嘌呤碱基U的偏好强于嘌呤碱基A,与本研究结果相似。本研究以竹节参转录组为数据来源,研究其密码子使用模式,确定竹节参表达基因的最优密码子,一方面可以利用密码子优化对外源基因进行密码子改造,提高外源基因在竹节参中的表达水平,同时也为竹节参相关基因的异源表达及分子育种等提供了理论依据。
参考文獻:
[1]Ikemura T. Codon usage and tRNA content in unicellular and multicellular organisms[J]. Molecular Biology and Evolution,1985,2(1):13-34.
[2]朱孝轩,朱英杰,宋经元,等. 基于全基因组和转录组分析的赤芝密码子使用偏好性比较研究[J]. 药学学报,2014(9):1340-1345.
[3]Fedorov A,Saxonov S,Gilbert W. Regularities of context-dependent codon bias in eukaryotic genes[J]. Nucleic Acids Research,2002,30(5):1192-1197.
[4]Hiraoka Y,Kawamata K,Haraguchi T A. Codon usage bias is correlated with gene expression levels in the fission yeast Schizosaccharomyces pombe[J]. Genes to Cells,2009,14(4):499-509.
[5]Quax T E,Claassens N J,Soell D,et al. Codon bias as a means to fine-tune gene expression[J]. Molecular Cell,2015,59(2):149-161.
[6]Zelasko S,Palaria A,Das A. Optimizations to achieve high-level expression of cytochrome P450 proteins using Escherichia coli expression systems[J]. Protein Expression and Purification,2013,92(1):77-87.
[7]李丽莎,李祥龙,周荣艳,等. 山羊酪氨酸相关蛋白1(TYRP1)基因密码子偏好性分析[J]. 贵州农业科学,2016,44(3):113-119.
[8]周宗梁,林智敏,耿丽丽,等. 水稻中cry1Ah1基因密码子优化方案的比较[J]. 生物工程学报,2012,28(10):1184-1194.
[9]李圣彦,郎志宏,朱 莉,等. 利用密码子优化提高Bt cry1Ah基因在转基因玉米(Zea mays L.)中的表达[J]. 中国农业科技导报,2011,13(6):20-26.
[10]杨金玲,高丽丽,朱 平,等. 蝎毒镇痛活性肽基因BmK AngM1的密码子优化及其真核表达分析[J]. 药学学报,2012(10):1389-1393.
[11]蒋 玮,吕贝贝,何建华,等. 草菇密码子偏好性分析[J]. 生物工程学报,2014,30(9):1424-1435.
[12]范三红,郭蔼光,单丽伟,等. 拟南芥基因密码子偏爱性分析[J]. 生物化学与生物物理进展,2003,30(2):221-225.
[13]李 滢,匡雪君,孙 超,等. 川贝母转录组密码子使用偏好性分析[J]. 中国中药杂志,2016,41(11):2055-2060.
[14]陈 哲,胡福初,王祥和,等. 菠萝转录组基因密码子使用偏好性分析[C]//中国热带作物学会2016年学术年会论文集,2016:2.
[15]张 来. 黔产竹节参种子萌发试验研究[J]. 种子,2012,31(5):75-78.
[16]罗正伟,张 来,吕翠萍,等. 竹节参离体培养及植株再生[J]. 中药材,2011,34(12):1818-1823.
[17]张 来,张显强,罗正伟,等. 竹节参毛状根培养体系的建立及人参皂苷Re的合成[J]. 中国中药杂志,2010,35(18):2383-2387.
[18]Zhang L,Sun M. Molecular cloning and sequences analysis of SS gene from Panax japonicus[J]. Research Journal of Biotechnology,2014,9(6):59-63.
[19]张 来,孙 敏. 贵州民间苗药竹节参叶的生药鉴定[J]. 中药材,2009,32(5):691-693.
[20]Meyer C A,Zhang L,Zhang X Q,et al. Comparative analysis of the essential oils from normal and hairy roots of Panax japonicas[J]. African Journal of Biotechnology,2011,10(13):497-519.
[21]张 来,杨碧昌,黄元射. 黔产竹节参根人参皂苷提取工艺与RP-HPLC含量分析[J]. 广东农业科学,2015,42(20):86-90.
[22]Rai A,Yamazaki M,Takahashi H,et al. RNA-seq transcriptome analysis of panax japonicus,and its comparison with other panax species to identify potential genes involved in the saponins biosynthesis[J]. Frontiers in Plant Science,2016,7(e0144):481.
[23]Zhang W J,Zhou J,Li Z F,et al. Comparative analysis of codon usage patterns among mitochondrion,chloroplast and nuclear genes in Triticum aestivum L.[J]. Journal of Integrative Plant Biology,2007,49(2):246-254.
[24]Fuglsang A. The effective number of codons for individual amino acids:some codons are more optimal than others[J]. Gene,2003,320(3):185-190.
[25]Comeron J M,Aguadé M. An evaluation of measures of synonymous codon usage bias[J]. Journal of Molecular Evolution,1998,47(3):268-274.
[26]Sharp P M,Li W H. The codon adaptation index—a measure of directional synonymous codon usage bias,and its potential applications[J]. Nucleic Acids Research,1987,15(3):1281-1295.
[27]Bellgard M,Schibeci D,Trifonov E,et al. Early detection of G+C differences in bacterial species inferred from the comparative analysis of the two completely sequenced helicobacter pylori,strains[J]. Journal of Molecular Evolution,2001,53(4/5):465.
[28]Kawabe A,Miyashita N T. Patterns of codon usage bias in three dicot and four monocot plant species[J]. Genes & Genetic Systems,2003,78(5):343.
[29]Saul A,Battistutta D. Codon usage in Plasmodium falciparum[J]. Molecular and Biochemical Parasitology,1988,27(1):35-42.
[30]Muto A,Yamao F,Osawa S. The genome of Mycoplasma capricolum[J]. Progress in Nucleic Acid Research & Molecular Biology,1987,34:29.
[31]Dybvig K,Voelker L L. Molecular biology of Mycoplasmas[J]. Annual Review of Microbiology,2003,50(50):25-57.
[32]劉庆坡,薛庆中. 粳稻叶绿体基因组的密码子用法[J]. 作物学报,2004,30(12):1220-1224.
[33]续 晨,贲爱玲,蔡晓宁. 蝴蝶兰叶绿体基因组密码子使用的相关分析[J]. 分子植物育种,2010,8(5):945-950.
[34]尚明照,刘 方,华金平,等. 陆地棉叶绿体基因组密码子使用偏性的分析[J]. 中国农业科学,2011,44(2):245-253.
[35]李秀璋,宋 辉,李春杰. 茄腐镰孢(Fusarium solani)线粒体基因组密码子偏好性分析[J]. 基因组学与应用生物学,2015,34(11):2465-2472.
[36]李 滢,匡雪君,朱孝轩,等. 长春花密码子使用偏好性分析[J]. 中国中药杂志,2016,41(22):4165-4168.
