小麦花粉育性检测和花粉致死基因携带种质的鉴定
2019-08-13
(西北农林科技大学农学院,国家杨凌农业生物技术育种中心/国家小麦改良中心杨凌分中心/小麦育种教育部工程研究中心/陕西省作物杂种优势研究与利用重点实验室, 陕西 杨凌 712100)
小麦(TriticumaestivumL.)是我国第二大粮食作物,其产量直接影响粮食平衡。因此提高小麦产量,降低小麦生产成本,提高国际竞争力成为我国国民经济和和社会发展课题中的一项重要内容。在生物界中,杂种优势是一种普遍现象,利用杂种优势实现作物的高产育种已在玉米、水稻等作物中取得显著成果。但是在利用作物杂种优势研究中发现,2个完全结实正常的品种杂交后,其F1代植株常常表现异常,如叶片坏死、植株黄化甚至枯萎、个体矮小、结实明显下降等症状。Sax[1]在研究小麦杂种的不育性时发现,Bluestem/Amby的正交和反交F1在苗期初期表现正常,当株高达到15~20 cm时植株生长停滞、过度分蘖,随后死亡。Loegering等[2]和Sears等[3]在普通小麦品种“中国春”和澳大利亚春小麦品种“Timstein”的杂交F1植株上发现,大约有50%花粉败育。在玉米中,ZmAA1基因被报道为花粉致死基因,编码了一个α-淀粉酶蛋白,该蛋白属于家族糖基水解酶,可催化水解多糖分子(1-4)-α-D-糖苷键,通过特异干扰花粉中淀粉的正常积累,从而抑制花粉正常发育最终导致花粉发育夭折[4-5]。Chang等[6]在水稻中发现OsNP1的隐性核雄性不育基因。OsNP1编码假定的葡萄糖-甲醇-胆碱氧化还原酶,该酶调节绒毡层的降解和花粉外壁形成;主要在绒毡层和小孢子中表达。osnp1突变体营养生长正常,但完全不育,并对环境条件不敏感。在小麦中,王鹏等[7]以中国春和澳大利亚春小麦品种的BC1F1代作为定位群体对花粉致死基因Ki进行分子标记定位,并将Ki基因定位于6 B染色体长臂上,但筛选得到的2个遗传标记距离较远,其可靠性有待验证。
本研究试图通过组配多个杂交组合,鉴定花粉败育情况,从而证明花粉致死遗传模式在冬小麦品种中也是一种普遍存在的现象,同时对携带花粉致死基因的种质进行鉴定,以期为进一步克隆小麦花粉致死基因奠定基础。
1 材料和方法
1.1 材 料
试验所用155个小麦杂交组合F1及其亲本,中国春(携带显性花粉致死基因Ki),宁春4号和宁春16号小麦品种(携带隐性花粉致死基因ki[7])均由西北农林科技大学农学院小麦生殖发育与杂优利用课题组提供。
1.2 方 法
1.2.1材料种植
试验在西北农林科技大学试验农场进行。2015年4月选取62个小麦品种(系)通过人工去雄、授粉共组配155个杂交组合。 10月将F0种植于试验地,每个组合种植2行,行长1 m,行距25 cm,株距10 cm。2016年4月在小麦开花期取所有组合成熟花药用1% KI-I2染色,鉴定花粉育性。2016年11月,将经过鉴定具有半不育花粉育性F1组合的亲本以及带有花粉致死基因的中国春、宁春4号、宁春16号种植于西北农林科技大学小麦中心温室,通过人工去雄、授粉配制组合,收取F0种子。2017年3月将春化后的F1种植于大田,5月在小麦开花期取成熟花药用1% KI-I2溶液染色鉴定花粉育性。
1.2.2KI-I2溶液的配制
取2 g KI溶于5~10 mL蒸馏水中,然后加入1 g I2,待全部溶解后,再加蒸馏水定容至300 mL。贮于棕色瓶中备用。
1.2.3小麦花粉育性的检测
采集小麦成熟花药置于干净的载玻片上,然后用镊子将花药充分捣碎,使花粉释放,然后在破碎组织上滴1~2滴1% KI-I2溶液,盖上盖玻片,用吸水纸吸掉多余溶液,随后置于光学显微镜下观察并拍照。观察3个穗子(来自不同植株),每个穗子观察3朵小花,每片取3个视野。以目测不育花粉粒约占总花粉粒的50%为标准,检测所有的F1组合。
2 结果与分析
2.1 花粉育性检测
在小麦开花期,取成熟花药用KI-I2溶液染色,对可育花粉、不育花粉以及半不育花粉进行镜检观察。可育花粉粒染色呈深蓝色圆形,花粉粒较大(图1 A)。不育花粉粒较小,不规则,且染不上色(图1 B)。半不育花粉中,一半的花粉粒呈深蓝色圆形,花粉粒较大;另外一半的花粉粒较小,不规则,且染不上色,呈现败育状态(图1 C)。
2.2 小麦杂交组合F1的花粉育性检测
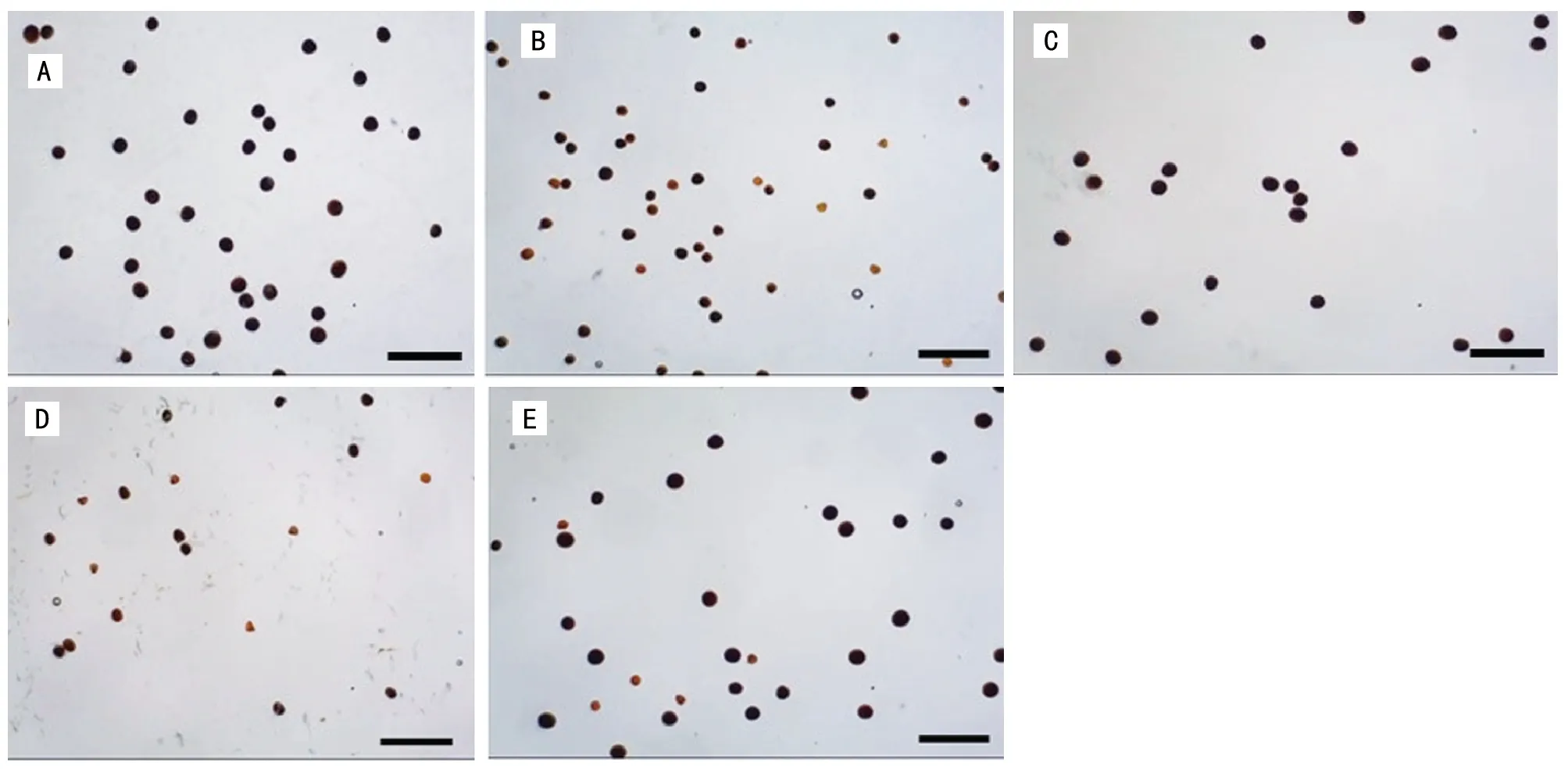
为了研究Loegering等[2]发现的春小麦花粉致死现象在普通冬小麦中是否也普遍存在,本研究选取了155个F1杂交组合,通过KI-I2溶液观察成熟花药。研究表明,大多数F1组合的花粉表现为可育花粉粒状态,呈深蓝色圆形,花粉粒较大。但是有4个F1组合的花粉表现为半不育花粉状态,一半花粉粒呈深蓝色圆形,花粉粒较大;另外一半的花粉粒较小,不规则,染不上色。它们分别是西农9718/鑫农516,西农9817/周麦22,西农9817/西农509,西农261/存麦4号(图2)。

注:A为西农9817/鑫农516;B为西农9817/周麦22;C为西农9817/西农509;D为西农2611/存麦4号。Bar=0.5 mm。图2 小麦杂交组合F1花粉活性
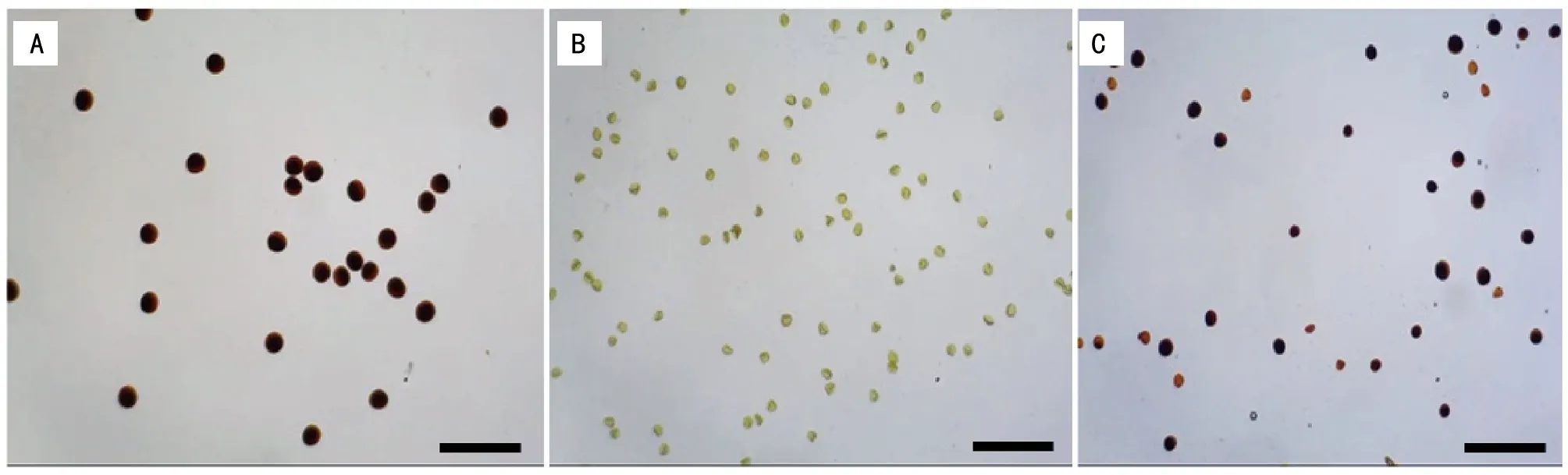
注:A为可育花粉;B为败育花粉;C为半不育花粉。Bar=0.5 mm。图1 KI-I2染色检验花粉活性
2.3 携带花粉致死基因品种及杂交组合的鉴定
Loegering等[2]Sears等[3]研究发现,小麦品种“中国春”的6 B染色体长臂上携带有显性花粉致死基因Ki,而澳大利亚春小麦品种“Timstein”携带有相应的隐性花粉致死等位基因ki,它们的杂交后代中,F1代出现了50%的花粉败育现象。王鹏等[7]利用“中国春”和“Timstein”鉴定出在普通春小麦品种宁春4号和宁春16号携带有隐性花粉致死等位基因ki。因此本研究以中国春和宁春4号、宁春16号与4个花粉表现均为半不育F1组合的亲本分别杂交获得相应F1,用KI-I2溶液为染料,对其成熟花药染色鉴定,从而鉴定携带花粉致死基因种质;鉴定标准为,对以中国春小麦品种为母本的杂交组合F1植株花粉育性进行检测,当其花粉不育率为50%时,则证明该组合相应父本品种携带等位隐性ki基因;对以宁春4号、宁春16小麦品种为母本的杂交组合F1植株花粉育性进行检测,当其花粉不育率为50%时,则证明该组合相应父本品种携带等位显性Ki基因。结果(图3,表1)表明,西农261和西农9817携带显性Ki基因,鑫农516,周麦22,西农509和存麦4号携带相应等位隐性ki基因。
注:A为存麦4号;B为西农2611/存麦4号;C为西农2611;D为存麦4号/CS;E为西农2611/宁春4号。Bar=0.5 mm。图3 携带花粉致死基因品种的花粉育性鉴定性
3 讨 论
保障粮食安全始终是我国农业科技现阶段的重要任务。目前,农作物育种技术中最广泛应用、最有效的技术之一就是杂交育种技术。杂交水稻、杂交油菜、杂交玉米等对世界粮食起着十分重要的贡献。杂交小麦的实现和应用,也必将对世界粮食供给起着巨大的作用。目前,已实现商业杂交种生产途径有细胞质雄性不育(CMS)系为基础的三系杂交、光温敏雄性不育(PTGMS)系为基础的两系杂交、化学杀雄(CHA)为基础的两系杂交、人工去雄为基础的两系杂交。尽管以上途径应用广泛,但是仍然存在一些固有的问题。CMS系统包括恢复系的种质资源狭窄,生产成本高、CMS系和恢复系之间的遗传差异较少等问题限制了CMS杂交育种的进一步发展[9];PTGMS系统种子的繁殖和杂交种子的生产,需要严格的环境条件,易受环境变化的影响[10];CHA系统则不可避免的涉及到环境污染的问题[11];人工去雄为基础的两系杂交,则难以降低种子生产成本。核雄性不育(包括显性和隐性)在开花植物中是很常见的,但是在杂种优势利用中一直无法实现,该类型不育系用于商业化生产杂交种子中的主要问题是难以获得稳定的雄性不育保持系[11]。Driscoll[13]则设想出XYZ体系生产杂种小麦,但未获成功。1988年,黄寿松等[14-16]首创出附加型蓝标小麦核雄性不育,将长穗偃麦草(Agropyronelongatum)4 E染色体附加到小麦核型雄性不育系上(该4 E染色体上携带有蓝色胚乳基因和育性恢复基因),从而选育出由蓝色胚乳性状标记的小麦核雄性不育、保持系,简称为蓝标型(或BM型)小麦雄性不育、保持系。该体系中,浅蓝粒种子植株自交结实后粒色分离为深蓝、浅蓝和白色3种,其中,占3%左右的深蓝色籽粒种子长出的植株自交结实,粒色为深蓝色,不分离;白色籽粒种子占64%左右,长出的植株全部表现为雄性不育,粒色不分离;而浅蓝色籽粒种子占33%左右,长出的植株仍然自交结实,粒色仍然分离为深蓝、浅蓝和白色3种。这样白粒种子长出雄性不育植株,可用来配制组合生产杂交种种子;而浅蓝粒种子的植株则可以用来进一步繁殖雄性不育系和保持系种子。1998年,李中安[17]创制了易位型蓝标小麦核雄性不育,将紧密连锁的蓝色胚乳基因和育性恢复基因易位到“Probus”核雄性不育系4 BS染色体上,杂交种生产原理同附加型蓝标小麦核雄性不育。这种杂交种生产体系解决了小麦核基因控制的雄性不育系在生产中难以获得稳定的保持系,难以获得大量纯合雄性不育系种子的问题。但是,在以浅蓝粒作为保持系的自交繁殖群体中,存在3%左右的深蓝粒,从而影响了不育系的繁殖效率。据此,王鹏等[7]提出将花粉致死基因ki转到4 E染色体上,从而蓝粒标记基因、雄性可育基因Ms和花粉致死基因ki被紧密连锁在一起,使4 E单价体无法通过雄配子传递,这样就能彻底解决深蓝粒的存在问题。
表1 携带花粉致死基因小麦种质鉴定
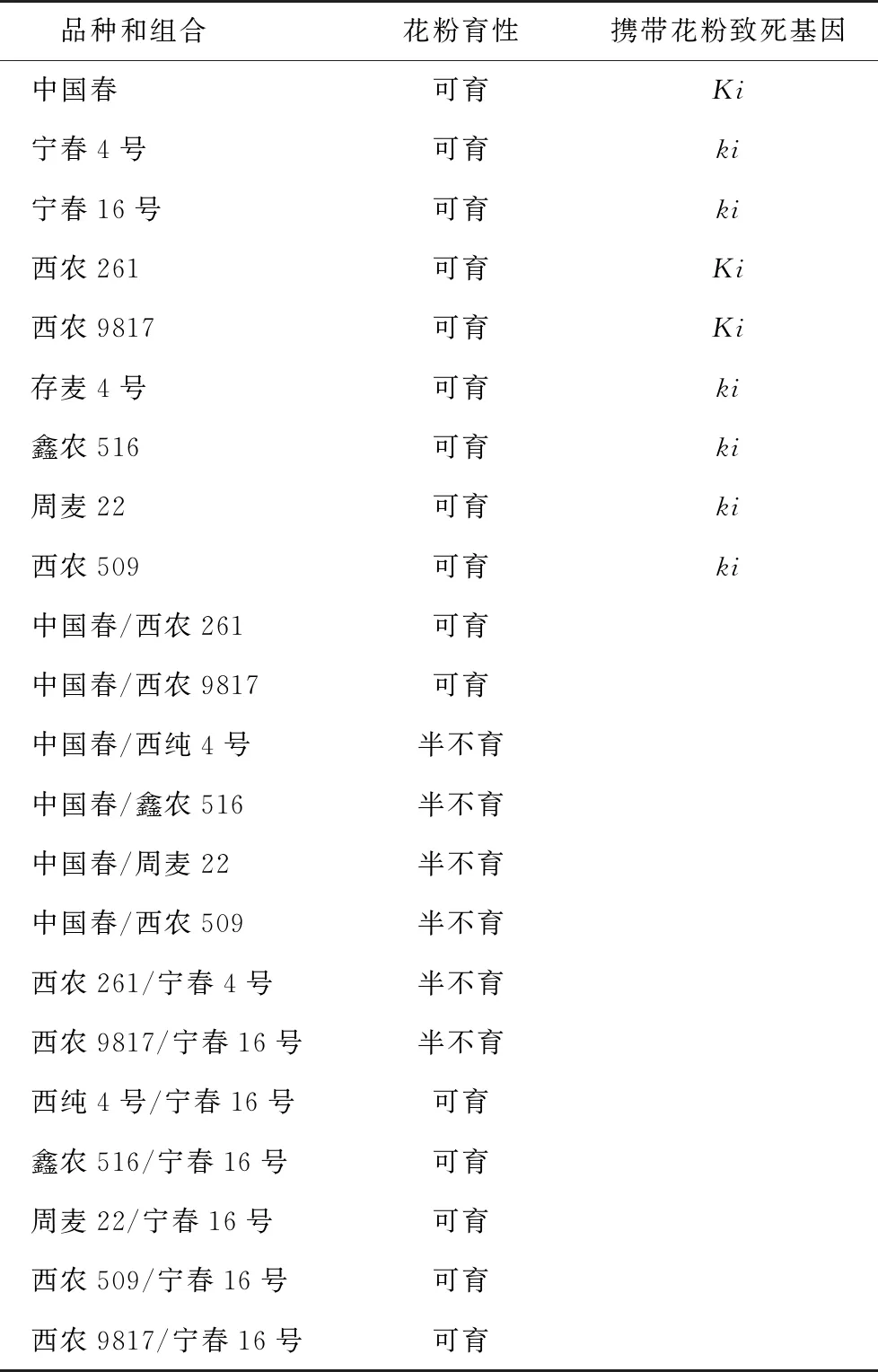
品种和组合花粉育性携带花粉致死基因中国春可育Ki宁春4号可育ki宁春16号可育ki西农261可育Ki西农9817可育Ki存麦4号可育ki鑫农516可育ki周麦22可育ki西农509可育ki中国春/西农261可育中国春/西农9817可育中国春/西纯4号半不育中国春/鑫农516半不育中国春/周麦22半不育中国春/西农509半不育西农261/宁春4号半不育西农9817/宁春16号半不育西纯4号/宁春16号可育鑫农516/宁春16号可育周麦22/宁春16号可育西农509/宁春16号可育西农9817/宁春16号可育
Loegering等[2]研究发现,“中国春”与澳大利亚春小麦品种“Timstein”杂交后,F1植株花粉不育率为1/2,由此提出小麦花粉致死基因的单基因遗传模式。Kato等[8]研究认为,小麦品种间杂交F1植株中,花粉不育率不是1/2,而是1/4或1/8,并且由此提出小麦花粉致死性状是由3个分别命名为Ki2,Ki3和Ki4的互补基因控制的。本研究通过组配155个F1杂交组合,在小麦开花期对成熟花药用1% KI-I2染色观察,证明这种花粉致死现象在普通小麦中是广泛存在的。并且分别以“中国春”、宁春4号、宁春16号春小麦品种为母本的杂交组合,对花粉致死基因做了遗传模式分析,发现不同组合杂交后,存在花粉异常的F1植株的花粉不育率都为1/2,该结果与之前Loegering和Sears等[2-3]研究结果一致,由此说明花粉致死基因在普通小麦品种中为单基因遗传模式[7]。
2009年美国先锋公司利用玉米SPT制种技术,构建了Ms 45-ZmAA1-DsRed 2基因盒的表达载体,对花粉致死基因ZmAA1及启动子Pg47应用其中。选种时通过光电筛选机利用DsRed红色荧光挑选种子,可满足全程机械操作且繁殖时只需隔离自交,成功解决了核雄性不育两系法运用中的问题[18]。利用花粉致死基因导致携带有外源育性基因的雄配子死亡,使转基因元件不会遗传给杂交后代,因此有效地解决了受到全球争议的转基因食品的安全性问题[19]。与上述技术类似,Chang等[6]利用水稻隐性核不育基因OsNP1与一个α-淀粉酶基因和红色荧光蛋白(DsRed)基因相结合,构建载体并转化osnp1突变体,这样携带单合子转基因水稻自交,可产生1∶1的比例雄性不育种子和转基因的可育种子,其中转基因种子可利用DsRed红色荧光挑选出来,雄性不育种子作为不育系可用来生产杂交种。这样的新型不育系兼具三系法的稳定性和两系法配组灵活性的优点,被称为 “第三代杂交育种技术”(G 3育种技术)[19]。
花粉致死基因在今后作物分子设计不育系技术研究中将起到十分重要的作用,突破作物杂交育种分子设计技术,创制出不受环境限制的、稳定的、具有恢复功能的新型不育系,实现作物杂种优势利用技术的新变革。
