裂拟迷孔菌生物学特性及驯化栽培分析*
2019-08-13郭艳芳
汪 阳,李 硕,张 桐,郭艳芳,李 丹,张 波,李 玉
(吉林农业大学食药用菌教育部工程研究中心,吉林 长春 130118)
木质纤维类生物质是自然界中最大的有机碳源库,白腐真菌可以分解此类物质,该类真菌在自然界碳循环中起着极其重要作用[1]。白腐真菌在分解木质素过程中会产生锰过氧化物酶(manganese peroxidase,MnP)、木质素过氧化物酶 (lignin peroxidase)、漆酶(laccase) 3种木质素酶氧化与分解木质素,使白腐真菌及其酶在将木材转化为食品、饲料或纤维制品方面具有潜在应用性。其中锰过氧化物酶不但可以使存在于木质素中的酚类结构单元被氧化,也可以使非酚类结构单元被氧化,是高效的木质素降解酶[2-3]。因为MnP具有降解木材中木质素的能力而被应用于造纸业,且MnP也可对有毒酚类物质染料等降解脱色,被应用于工业废水的处理中,所以近年对MnP的研究得到了广泛关注[4]。
裂拟迷孔菌(Daedaleopsis confragosa),俗名粗糙拟迷孔菌,隶属于真菌界(Fungi)担子菌门(Basidiomycota) 蘑菇亚门 (Agaricomycotina) 蘑菇纲 (Agaricomycetes) 多孔菌目 (Polyporales) 多孔菌科 (Ployporaceae) 拟迷孔菌属 (Daedaleopsis)[5]。该种广泛分布在北温带林地中,常生长在柳树群落(湿地或沼泽)的老树上,也可生长在其他阔叶树木上。该种英文名字(blushing bracket)源于其受伤后的变红反应,还有一个可替代的英文名字(thinwalled maze polypore)[6]。
裂拟迷孔菌子实体木栓质,菌盖扇形至半圆形,菌盖表面浅黄色至褐色,幼时被细弱绒毛,后期变光滑,具同心环带,边缘较锐。孔口表面乳黄色,近圆形、长方形、迷宫状;担孢子(6.1~7.8) μm×(1.2~1.9) μm,圆柱形,无色,光滑,非淀粉质,不嗜蓝[5,7]。
裂拟迷孔菌多应用在东方造纸术中,子实体干燥后可做成纸浆制成有纹理和颜色的纸张。其造成木材白色腐朽,具有木质素酶活性。但国内对其菌丝活性和驯化栽培研究未见报道,本试验对该菌菌丝和驯化栽培以及锰过氧化物酶活性进行了研究分析,以期为该菌的进一步开发应用提供参考。
1 材料方法
1.1 试验材料
菌种:野生子实体采自辽宁省白狼山国家级自然保护区,采集人高洋,标本号41558,现藏于吉林农业大学菌种保藏中心(CCMJ),菌种编号2790。通过PDA培养基进行菌种分离培养,置于4℃恒温保存。
1.2 供试培养基
PDA:马铃薯200 g、葡萄糖20 g、琼脂18 g,蒸馏水1 000 mL,pH不做处理[8];碳源培养基配方:碳源20 g、磷酸二氢钾3 g、酵母膏2 g、蛋白胨2 g、硫酸镁1.5 g、琼脂18 g,蒸馏水1 000 mL,pH不作处理;pH培养基配方:马铃薯200 g、葡萄糖20 g、硫酸镁1.5 g、磷酸二氢钾3 g、琼脂18 g,蒸馏水1 000 mL;温度培养基配方:普通PDA培养基[9-10]。
1.3 试验方法
对碳源培养基作4个处理:葡萄糖、蔗糖、可溶性淀粉、糊精,对照组不添加碳源;氮源培养基作4个处理:蛋白胨、酵母浸粉、磷酸氢二铵、硝酸铵,对照组不添加氮源;pH作5个处理:5、6、7、8、9(灭菌后用1 mol·L-1盐酸和氢氧化钠调节);温度作6个处理:15℃、20℃、25℃、30℃、35℃、40℃,每个因素、每个处理各3次重复。选用7 mm规格打孔器打孔接种至培养皿,黑暗培养,十字交叉法测量菌丝生长速度,间隔24 h划线[11-12]。
正交试验培养基:碳源20 g、氮源2 g、磷酸二氢钾2 g、硫酸镁1 g、琼脂粉20 g,蒸馏水定容至1 000 mL,调节相应的温度和pH。
MnP活力测定:无菌条件下,从已活化的菌株中选取直径7 mm接种块5个,接种至50 mL液体发酵培养基中,摇床30℃、150 r·min-1条件下震荡培养。自接种后24 h开始,每间隔24 h取1.0 mL菌液于1.5 mL无菌离心管中,-4℃低温离心后取上清液,-20℃保存。11 d后,统一使用试剂盒(苏州科铭生物技术有限公司)进行MnP活力测定,试验重复3次。
驯化栽培:从二级种制备、栽培出菇措施方面进行分析。
2 结果与分析
2.1 鉴定
使用新型植物基因组DNA提取试剂盒(康为世纪) 提取DNA,选用通用引物ITS1和ITS4对裂拟迷孔菌进行PCR扩增,将获得的自测序列(WY2790)在NCBI数据库进行比对,与X-78序列具有98%的相似性,定其为裂拟迷孔菌Daedaleopsis confragosa。选取NCBI数据库序列以多孔菌属Polyporus作为外群,使用Mega 7软件以最大似然法构建系统发育树,Daedaleopsis聚类到一支,另一支为外群Polyporus,如图1所示。
从图1可以看出,自测序列(WY2790) 与NCBI中同种序列聚集在一起,可以辅助证明,该种正是裂拟迷孔菌Daedaleopsis confragosa。
2.2 不同碳源对菌丝生长速度影响比较
4种碳源对菌丝生长的影响见表1。
由表1得出,裂拟迷孔菌菌丝在可溶性淀粉、糊精2种碳源培养基上生长较好,在蔗糖、葡萄糖2种碳源培养基上生长较慢,虽然菌丝以可溶性淀粉和糊精为碳源时,生长速度差异并不明显,但糊精培养基中菌落边缘不整齐,故总体来看,可溶性淀粉是最佳碳源。其次葡萄糖培养基中裂拟迷孔菌菌落边缘整齐,蔗糖培养基菌落边缘不整齐且菌丝生长速度最慢。从菌丝长势看,裂拟迷孔菌菌丝在4种碳源培养基中菌丝都较为浓密。综合菌丝生长速度、菌丝长势,裂拟迷孔菌菌丝对供试4种碳源的喜好顺序为可溶性淀粉>糊精>葡萄糖>蔗糖。
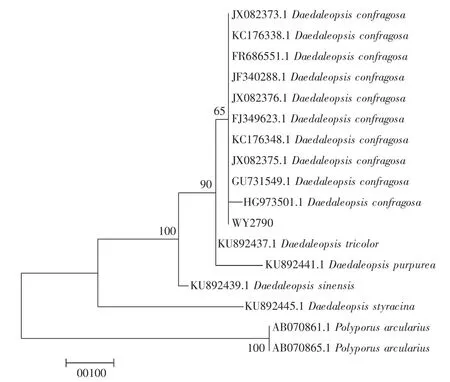
图1 最大似然法构建的基于裂拟迷孔菌属与多孔菌属ITS序列的系统发育树Fig.1 A phylogenetic tree based on the ITS sequence of the genus Daedaleopsis and the genus Polyporus constructed by the maximum likelihood method
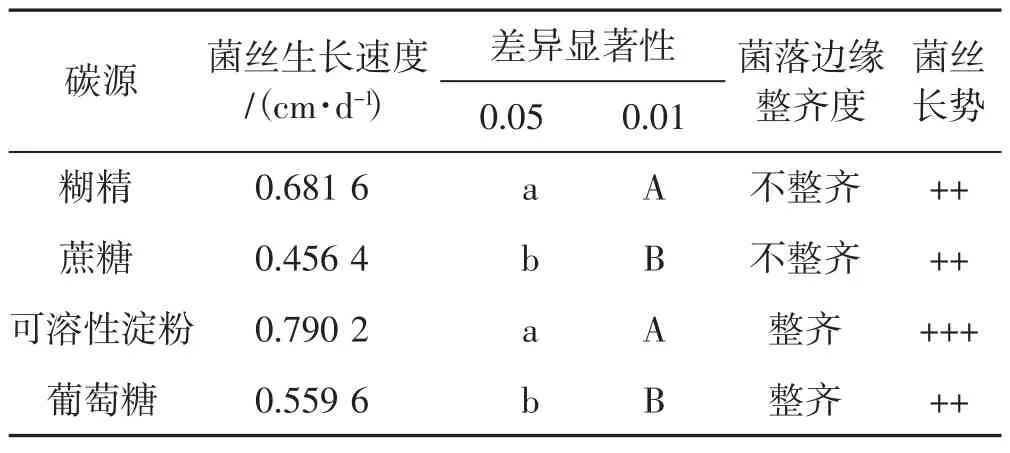
表1 4种碳源下菌丝生长情况Tab.1 Mycelial growth with four carbon sources
2.3 不同氮源对菌丝生长速度影响比较
4种氮源对菌丝生长速度影响见表2。
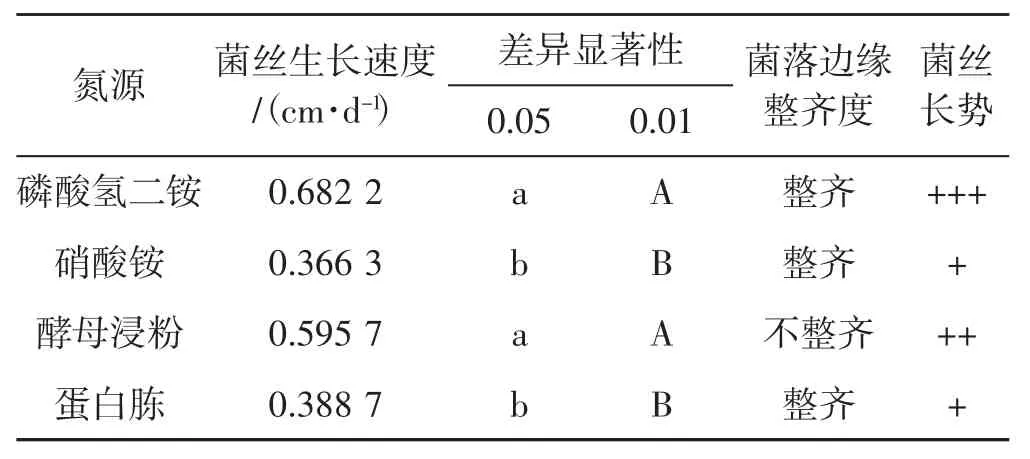
表2 4种氮源下菌丝生长情况Tab.2 Mycelial growth with four nitrogen sources
从表2可以看出,裂拟迷孔菌菌丝在磷酸氢二铵、酵母浸粉2种氮源源培养基上生长较好,在蛋白胨、硝酸铵2种氮源培养基上生长较慢。酵母浸粉培养基中菌落边缘生长极不整齐,故磷酸氢二铵是最佳的供试氮源。而以蛋白胨、硝酸铵作为培养基时,菌丝生长速度明显减弱。综合菌丝生长速度和菌丝长势可以得出,菌丝对4种供试氮源适应性差异顺序为磷酸氢二铵>酵母浸粉>蛋白胨>硝酸铵。
2.4 不同pH值对菌丝生长速度影响比较
5个pH值对菌丝生长速度影响见表3。
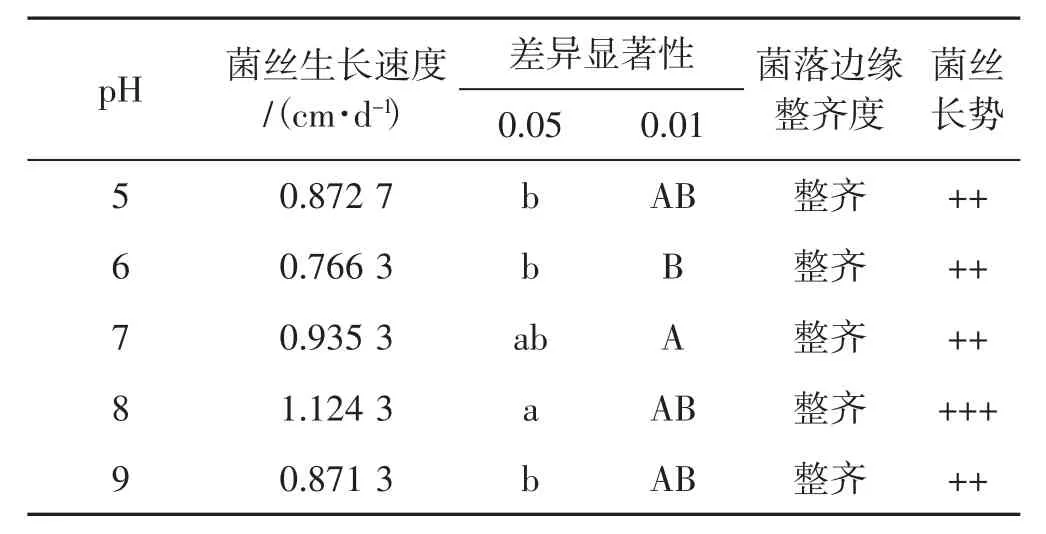
表3 5个pH值下菌丝生长情况Tab.3 Mycelial growth with five pH values
由表3得出,裂拟迷孔菌菌丝在pH为8时生长最好,其他pH时生长差异并不显著。从菌丝生长速度得出,在5个不同pH情况下,菌丝长势并无明显差别,pH值为5和9时菌丝生长速度差异不明显,pH值为6时菌丝生长速度最慢。综合菌丝生长速度、长势得出,裂拟迷孔菌菌丝不同pH生长优劣情况为pH8>pH7>pH5>pH9>pH6。
2.5 不同温度梯度对菌丝生长速度影响比较
6个温度梯度对菌丝生长速度影响见表4。
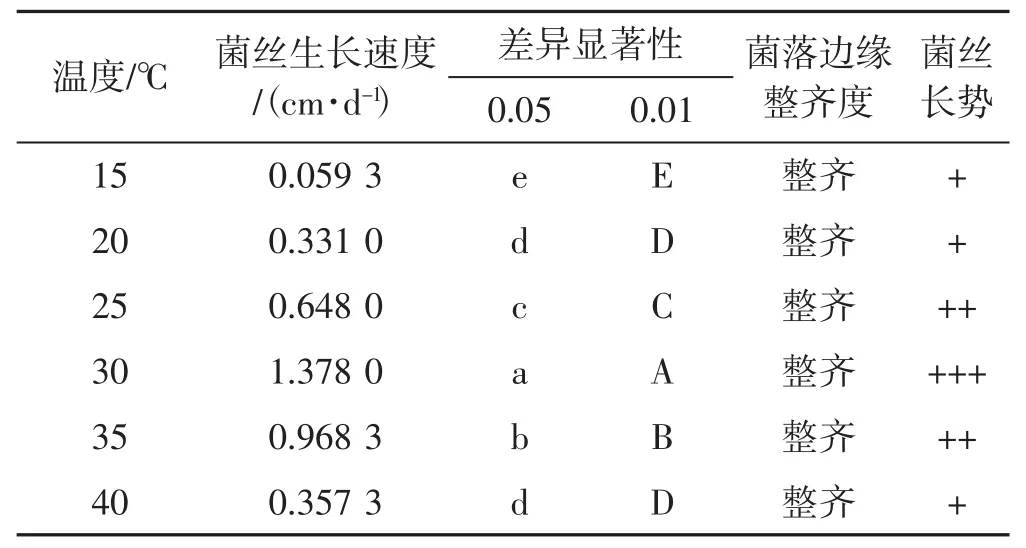
表4 6个温度梯度下菌丝生长情况Tab.4 Mycelial growth with 6 temperature gradients
如表4所示,裂拟迷孔菌菌丝菌落直径从起初接种1 d后至试验结束,30℃下的培养皿菌落皆保持着生长最优势。裂拟迷孔菌平板菌丝在15℃到30℃之间,菌丝生长情况与温度升高成正比。裂拟迷孔菌菌丝对温度的生长喜好为30℃>35℃>25℃>40℃>20℃>15℃。
2.6 正交试验
单因素试验并未考虑到各个单因素间交互作用的影响,不能将其结果作为最终结果。因此对氮源、碳源、pH和温度4个因素进行正交试验,从以上试验中挑出每种因素3个最优水平,组建4因素3水平的正交分析表(表5),以期选出最适宜的搭配组合,为裂拟迷孔菌的广泛应用奠定基础。菌丝生长正交试验结果直观分析和方差分析见表6~表7。

表5 L9(34)正交试验因素水平Tab.5 Factor level of L9(34)orthogonal test
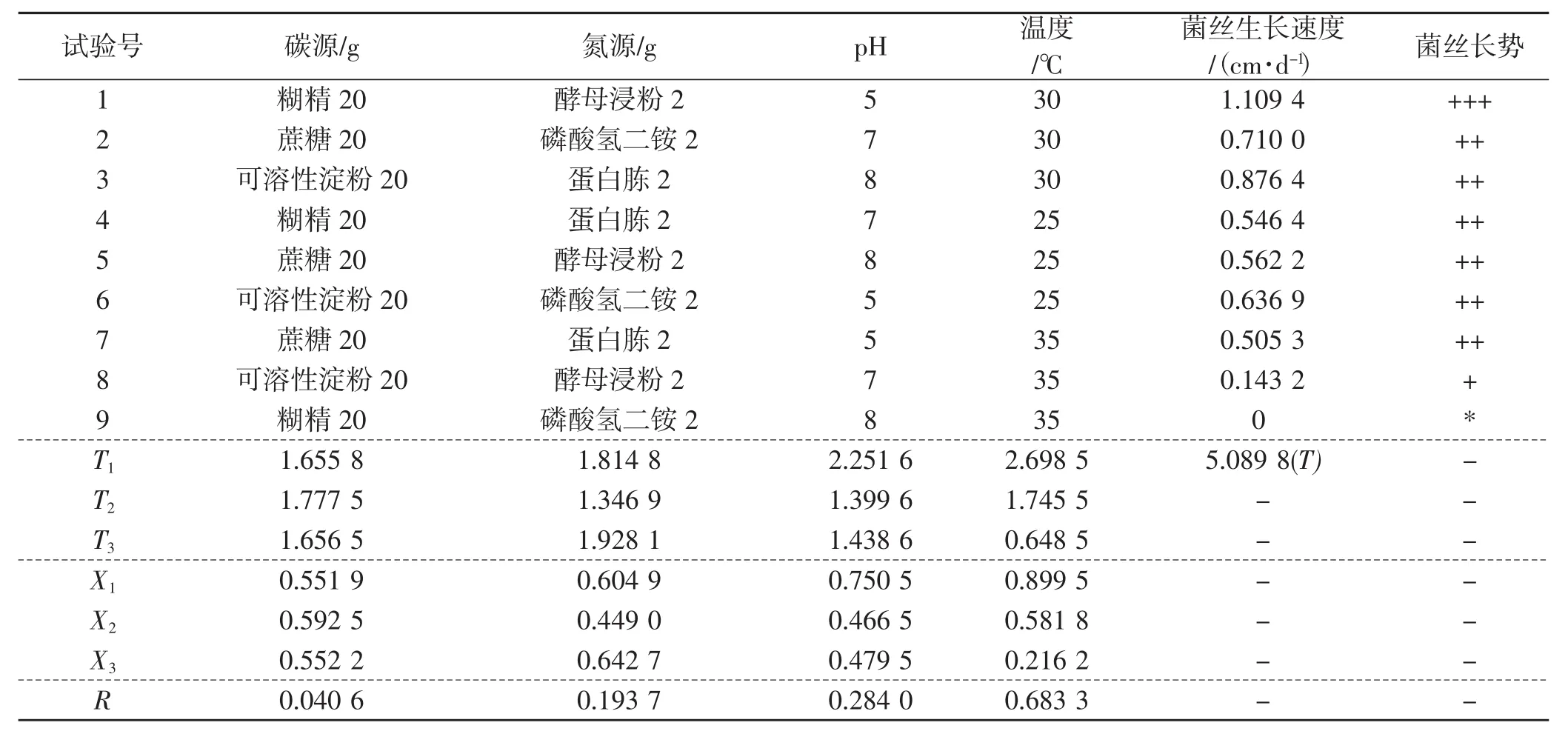
表6 菌丝生长正交试验结果直观分析Tab.6 Visual analysis of orthogonal test results of mycelial growth
从表6正交试验结果直观分析得出,温度是影响裂拟迷孔菌最主要的因素,其极差达到0.683 3。然后是pH,极差为0.284。碳源、氮源的极差分别为0.040 6、0.193 7。从表6中可以看出,碳源的均值大小顺序为X2>X3>X1,所以选择蔗糖(X2),依次类推氮源选择蛋白胨(X3),pH选择pH=5(X1),温度选择30℃(X1)。
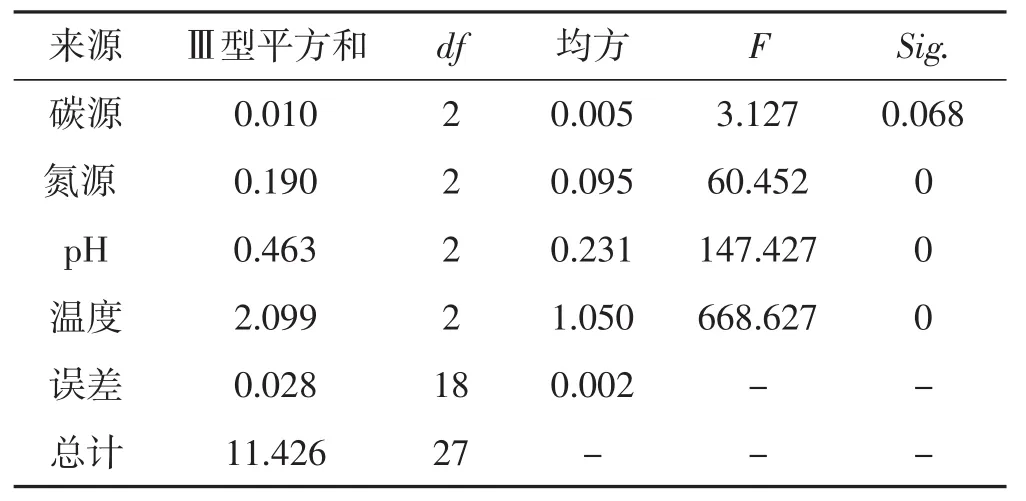
表7 菌丝生长正交试验结果方差分析Tab.7 Analysis of variance of orthogonal test results of mycelial growth
从表7可以看出,4种生长因素对裂拟迷孔菌菌丝生长影响显著性差异顺序是温度>pH>氮源>碳源,与直观分析结果一致。
2.7 锰过氧化物酶活力测定
以正交试验最优组合并加入硫酸镁和磷酸二氢钾作为液体发酵培养基,锰过氧化物酶活力测定结果见图2。

图2 裂拟迷孔菌在液体发酵培养基中MnP活力Fig.2 Manganese peroxidase activity of Daedaleopsis confragosa in liquid fermentation medium
结果显示MnP活力在第1天至第6天活力低且波动不剧烈,第7天活力开始上升,第10天出现峰值,达到19.4110U·L-1,第11天急剧下降为0.1652U·L-1。
2.8 驯化栽培
二级种选用麦粒菌种,500 mL罐头瓶装干料160 g,每瓶接整个培养皿接种块,25℃培养[13]。
三级种配方:阔叶树木屑78%、麸皮20%、石膏1%、蔗糖1%,加水搅拌充分混匀培养料,湿度约65%,装入规格为17 cm×34 cm的菌袋中,每个菌袋装湿料700 g,中间插入孔径1.5 cm的打孔棒。121℃高压灭菌2 h,无菌条件冷却至25℃以下接种。
发菌和后熟:将已接入二级种的菌袋放入发菌室,保证室内温度18℃~22℃,湿度60%~65%,黑暗培养。裂拟迷孔菌菌丝在28 d长满菌袋。菌袋继续放在发菌室后熟并严格控制发菌室温度、湿度,9 d后完成后熟。
催蕾出菇:后熟完成后使出菇房温度降低至14℃~16℃,并在菌袋上割口。低温刺激有助于菇蕾的形成,在低温刺激11 d后原基形成。在出菇期间,应保持较高的空气湿度(95%左右),温度控制在23℃~25℃,促进子实体形成。
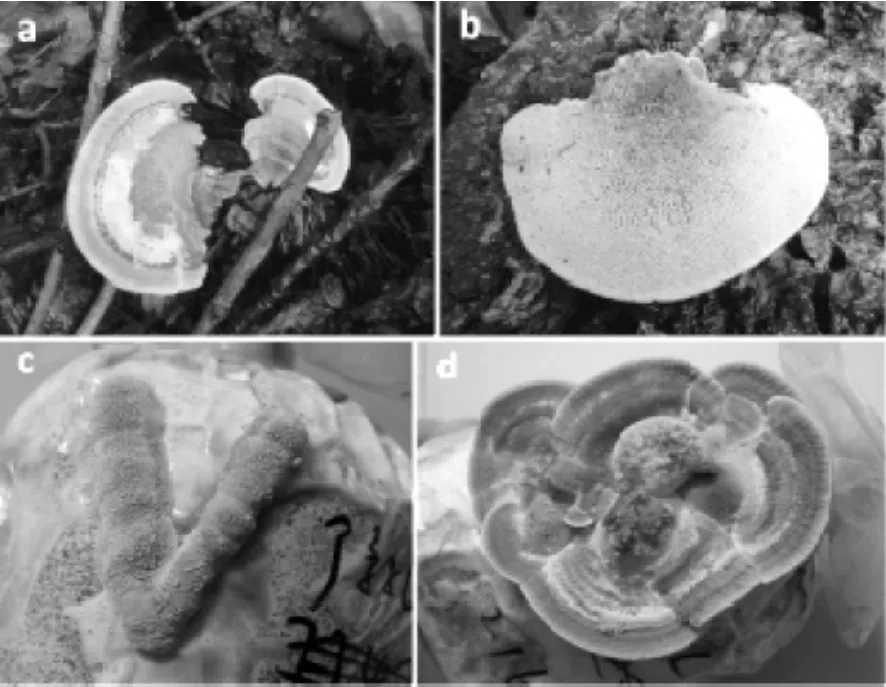
图3 裂拟迷孔菌野生子实体及驯化栽培子实体照片Fig.3 Photos of Daedaleopsis confragosa wild fruiting body and domesticated cultivated fruiting body.
3 结论与讨论
通过碳源、氮源、pH、温度单因素试验对裂拟迷孔菌菌丝生长的影响和正交试验结果,最终确定裂拟迷孔菌在糊精作为碳源,酵母浸粉作为氮源,pH为5,培养温度为30℃时长势最好。在碳源单因素试验中,菌丝对多糖碳源具有明显的偏好性,碳源单因素试验中可溶性淀粉培养基菌丝长势最好,与Gary[3]报道的可溶性淀粉是最易被真菌分解的多糖结果相同。在正交试验中,以糊精作为碳源时菌丝生长最优,其次可溶性淀粉,与单因素试验结果相反。正交试验考虑了各因素间的交互作用,前文已得出4种因素对裂拟迷孔菌菌丝生长影响显著性差异顺序是温度>pH>氮源>碳源,碳源较其他3种因素对菌丝生长优劣的影响较小,结合鲁铁[9]、丁野[10]、程国辉[14]单因素试验及正交试验结果推测,多种因素共同作用时菌丝的生长会偏重对其影响更大的因素,而对影响较小的因素采取一定适应性。
在氮源单因素试验中,菌丝长势较好的依次是无机氮源磷酸氢二铵和有机氮源酵母浸粉,正交处理中菌丝长势在酵母浸粉作为氮源时最优。蛋白胨和酵母浸粉都是有机氮源,而前者作为氮源时菌丝生长速度不及后者,这是因为酵母浸粉中氮源物质主要以易吸收的蛋白质降解产物形式存在,而降解产物特别是氨基酸可以通过转氨作用直接被菌丝利用,为速效氮源,有利于被菌丝更快地利用;蛋白胨中氮源物质存在的形式多是大分子蛋白质,要被菌丝吸收利用则需要进一步降解成小分子的肽和氨基酸,是迟效氮源,利用速度缓慢。将此最优的正交处理与正交分析表试验号2的正交处理相比,按前文所得的温度、pH是影响菌丝生长最重要的2种因素,且试验号2的正交处理选用氮源是单因素试验中最优氮源,可2种正交处理菌丝生长速度差异却接近0.4 cm·d-1,值得注意的是蔗糖为双糖,糊精为多糖,推测在复合处理时,多糖碳源相比其他碳源更能促进菌丝生长,这与Paul Ander&Karl-erik Eriksson[15]在研究中发现多糖物质的存在,对于白腐真菌菌丝生长特性的选择有较大影响一致。
本试验中,菌丝在30℃情况下生长最快,在35℃时,平板生长显著减慢,而接种块在40℃且菌丝长度达到约2 cm时,菌丝生长即停止。与刘秀明等[16]基于白灵侧耳菌丝保护酶活性与不同温度间的联系研究,推论温度过高会抑制菌丝酶活性,从而抑制菌丝的生长速度一致。
试验得出裂拟迷孔菌菌丝在pH值5~9范围内均长势较好,并且菌丝在pH为8的情况下生长最快,这与Sérgio Luiz Moreira Neto等研究得出大多数多孔菌适宜在pH显酸性环境中生长不一致[17]。虽然裂拟迷孔菌可以侵染活木,是森林木材病原菌,可其只在较少情况下才会侵染活木[18-19],而不同的酸碱环境可能是真菌对植物致病性的重要决定因素[20],Nir Yakoby等[21]、Dov Prusk y等[22]对 Sclerotinia sclerotiorum的研究中发现在酸性条件下,2种可能在感染植物过程中起重要作用的酶:acp1(编码非天冬氨酸蛋白酶)和pg1(编码内聚半乳糖醛酸酶)的表达活性升高,推测裂拟迷孔菌菌丝偏好碱性,可能是其较少生长于活立木上的原因。
裂拟迷孔菌为白腐真菌,目前可有效对木质素进行降解的是白腐真菌[23],试验得出的裂拟迷孔菌锰过氧化物酶活性峰值是19.411 U·L-1。作为一种诱导酶类,锰过氧化物酶的合成水平也特别易受诱导剂的影响。在何平等[4]对乳白耙齿菌的诱导剂筛选研究中,适宜的诱导剂能大幅度地提高乳白耙齿菌的MnP活性,使之从12.700 U·L-1升高到203.226 U·L-1。由于试验已得出裂拟迷孔菌可驯化栽培,后续试验可对裂拟迷孔菌进行诱导剂筛选,选出最优诱导剂,使其在染料等有毒酚类物质降解脱色与工业废水治理中发挥较好作用。同时裂拟迷孔菌可以有效地清除·OH[24]。由此可见,裂拟迷孔菌具有良好的经济应用前景。本试验初步探索了裂拟迷孔菌的生长条件,成功栽培出白腐真菌裂拟迷孔菌,为规模化的木质素降解的进一步开发、生产提供了基础数据,也对该属其他种的生物学特性驯化栽培研究提供了参考。后续试验可对裂拟迷孔菌的栽培配方、药理活性方面进行筛选研究,也可进一步深入探讨分解木质素酶系统等。
