吉林蛟河阔叶红松林中红松的空间格局分析
2019-07-30丁胜建
丁胜建
(辽宁省林业调查规划监测院,辽宁沈阳110122)
阔叶红松林主要分布在我国东北地区,是温带针阔混交林的典型代表。 在阔叶红松林中,红松是珍贵的乡土树种,是群落的支撑性树种。 20 世纪以来,随着我国经济的快速发展,原始阔叶红松林资源遭到严重破坏,形成大面积次生林群落。 1999年8月4日国务院将红松定为国家二级重点保护野生植物。 2013年红松被列入 《世界自然保护联盟》(IUCN)濒危物种红色名录。
植物种群分布格局是森林群落中种子扩散、种内和种间竞争、干扰、环境异质性等相互作用的最直接反映[1],分析物种的空间格局有助于认识该格局形成的生态学过程、种群的生物学特性及其与环境因子之间的相互关系[2]。 种群的空间分布格局通常有随机分布、聚集分布和均匀分布3 种状态[3]。 传统的空间格局分析方法,如检验、方差区组分析、最近邻距离分析等[4],只能分析单一尺度空间分布格局。 本文应用了二阶统计方法Wiegand-Moloney’s O-ring 函数进行分析,克服了传统方法分析尺度单一缺点。
1 研究区概况
样地位于吉林省吉林市蛟河林业实验区管理局,海拔890 m,地理坐标为东经127°35′-127°51′,北纬43°51′-44°05′,属长白山系张广才岭山脉。 该区属大陆季风性气候,年平均气温为3.5 ℃,年平均降水量在700~800 mm 之间, 土壤为山地暗棕色森林土,土层厚度20~80 cm。 主要树种有红松(Pinus koraiensis)、 胡桃楸(Juglans mandshurica)、 鱼鳞松(Picea jezoensis)、紫椴(Tilia amurensis)、沙松(Abies holophylla)、 色木槭 (Acer mono)、 臭松(Abies nephrolepis)、白牛槭(Acer mandshurica)、枫桦(Betula costata)、千金榆(Carpinus cordata)等。 主要下木有暴马丁香(Syringa reticulata var. mandshurica)、东北山梅花 (Philadelphus schrenkii)、 毛榛子(Corylus mandshurica)、刺五加(Eleuthercoccu senticosus) 、东北溲苏 (Deutzia amurensis) 等。 主要草本有苔草(Carex spp.)、山茄子(Brachybotrys paridiformis) 、蚊子草(Filipendula sp.)、小叶芹(Aegopodum alpestre)和蕨类(Adiantum spp.)等[5]。
2 研究方法
2.1 样地调查
建立30 hm2(500 m×600 m) 固定样地,首先划分为750 个20 m×20 m 的连续样方。再将20 m×20 m 样方划分为16 个5 m×5 m 的小样方。 对样地内胸径(DBH)≥1 cm 的乔木进行编号,并调查树种、胸径、树高、冠幅、枝下高及其所处坐标。
以往的研究表明, 阔叶红松林树高>20m 左右时,属主林层;树高在10~20m 之间属次林层;树高<10m 属林下层[6]。 根据邵国凡[7]等关于树高与胸径关系的方程, 当红松的胸径为30cm 时, 树高约为20m;当两者的胸径为10cm 时,树高约为10m.因此把红松分为DBH<10cm、10cm≤DBH<30cm、DBH≥30cm 3 个径级,分别代表主林层、次林层和林下层3 个不同的林层。
2.2 分析方法
在植物空间格局分析中,二阶统计方法Ripley’s K 函数得到了普遍应用, 但是Ripley’s K 函数分析结果具有累积性, 大尺度会累积小尺度的信息,因此会使分析尺度混淆[8-10]。 Wiegand-Moloney’s Oring 函数用半径为r, 宽度为w 的圆环替代了Ripley’s K 函数计算中所使用的半径为r 的圆,解决了尺度混淆的弊病[10、11]。
O-ring 单变量统计函数被用于分析单个对象的分布格局,用O11(r)表示统计值,O-ring 双变量统计函数被用于分析两个对象的格局,用O12(r)表示统计值。 双变量O-ring 的统计值:
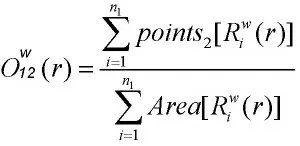
式中,n1为格局1 的点的数目,表示以格局1 中第i 个点为圆心,r 为半径,w 为宽度的圆环中包括的格局2 的点的数目。 表示研究区域的面积。 在单变量O-ring 分析中,假设格局1 的点和格局2 的点相同。
在本研究中,单变量O-ring 统计方法用于分析红松在3 个林层的空间分布格局,而双变量O-ring统计方法用于分析3 个林层间关联性。 数据分析过程使用生态学软件Programita (Wiegand,2006 版)完成。 采用的空间尺度为0~50m,19 次Monte Carlo 模拟得到95%的置信区间。
3 结果与分析
3.1 径级分布
在样地内共调查到DBH≥1cm 的红松1370株。 由图1 可以看出,红松的径级分布近似于L 型,小径阶的多,大径阶少。
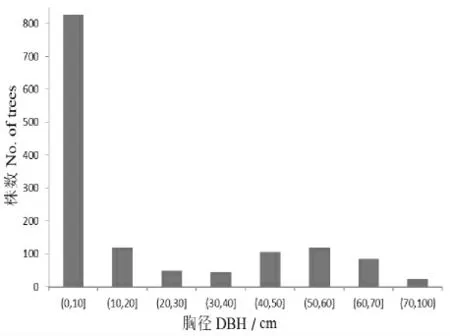
图1 红松与紫椴的径级结构Figure1 DBH class of P. koraiensis and T. amurensis
3.2 空间格局
对红松在不同林层的空间格局分析可以看出,林下层红松在0~50m 的尺度上均表现为聚集性分布,且在50 个的尺度上聚集性均较显著。 次林层红松在13m、15m、22m 尺度级上呈现出随机分布,其他47 个尺度级上均为聚集分布。主林层红松在2m、17m、20m、30m、38m、42-44m、46m、49m 尺度级上呈现随机分布,其他40 个尺度级均为聚集性分布(图2)。随着林层的升高,红松种群呈现出聚集性分布的尺度级数量在减少,且出现随机分布状态的起始尺度值在变小。
3.3 空间关联
红松种群的主林层与次林层在2-3m、11-12m、26m、38m 的尺度上表现出正相关,表明两个林层的个体之间具有相似的生态位,可以生长在相同的环境中;在40m 的尺度级上表现为负相关,只在这一尺度上不能说明红松主林层对次林层有抑制作用;而在其他尺度上,两个林层间关联性不显著(图3)表明两林层间个体可以共居一处,互不影响,保持相对独立。

图2 红松种群不同林层空间分布格局Figure 2 Spatial patterns of P. koraiensis at different vertical layers.
主林层与林下层2-18m、20-26m、28-36m、38-39m、44m、50m 的尺度上表现为显著的负相关,表明主林层红松对林下层红松有强烈的抑制作用;而在其他的尺度上,两个林层间关联性不显著(图3)表明两林层间个体共居一处,互不影响,保持相对独立。
次林层与林下层间在所检验的所有尺度上都显示出显著的正相关(图3),表明两个林层间存在相似的生态位和次林层对林下层有正的促进作用。

图3 红松种群不同林层空间关联Figure 3 Spatial associations of P. koraiensis at different vertical layers.
4 结论与讨论
通过计算红松在不同林层的O-ring 指数,结果显示,红松在所有林层都显示出了聚集性分布的特征, 但是随着尺度的增加逐渐表现为随机分布,且随着林层的升高,呈现出随机分布起始尺度值逐渐变小。 与以前的研究结果相一致[4、11],表明老龄阔叶红松林中,红松在较低林层比较高林层有强的聚集性。 红松种群在林下层表现出较强聚集性,主要原因受限于红松种子的传播方式。 红松种子的传播主要通过动物和种子自身的重力作用,红松种子不仅体积大且质量重,所以传播距离较短。 其次小径级个体需要的资源较少、竞争相对较弱,也导致低林层的红松有相对较强的聚集性。 红松种群在次林层逐渐表现出随机分布的状态, 说明随着红松的生长,个体间竞争逐渐增强。 在这个阶段红松种群内激烈的竞争,相互之间争夺光、水分和养分等资源,以满足其生存需要,导致了大量树木的死亡,即自疏效应。 种群内的自疏效应,使得红松种群在次林层的个别尺度级上表现出随机分布的状态。 红松种群在主林层相较次林层,在更多的尺度级上出现随机分布的状态,说明在主林层种群内依然存在激烈的竞争,自疏效应依然存在。
对红松种群不同林层的空间关联进行了分析,结果显示,红松群落的主林层与次林层在6 个尺度上表现为正相关,43 个尺度上表现为无显著关联,表明了红松具有耐阴特性,可以在自己的林冠下很好地存活和生长及次林层红松对环境适的应范围较宽。 主林层与林下层在37 个尺度级上表现为显著负相关,13 个尺度上表现为无显著关联, 说明红松在主林层占有了绝大多数环境资源,林下层处于绝对劣势地位。 红松作为群落的优势树种,红松在主林层排斥林下层红松的生长,从而促使红松幼树向未被红松主林层占领的区域发展,经过若干年生长演替主林层红松将占据整个群落。 红松群落的林下层和次林层在50 个尺度级上均表现出显著正相关,由于红松种子很难远距离传播,造成次林层红松种子多散落在母树附近,再有次林层红松相较主林层红松对环境资源的占有能力弱,保障了林下层红松生长发育所需资源和空间。
因此可以得出,红松种子扩散对其空间格局的形成起着决定性作用。 同时,随着红松个体间对生存资源的竞争的增强,自疏效应使得红松分布的聚集性下降。 当然可能还有其他方面的原因,例如一些未涉及的环境因子、干扰事件或随机因子的影响等。
