青稞 HVA1和 blt4.9基因对模拟水分胁迫的响应差异及其在抗旱育种中的应用
2019-07-24姚晓华吴昆仑白羿雄魏晓星
王 越,姚晓华,吴昆仑,白羿雄,魏晓星
(1.青海大学,青海西宁 810016; 2.青海省青稞遗传育种重点实验室/国家麦类改良中心青海青稞分中心,青海西宁 810016; 3.青海大学生态环境工程学院,青海西宁 810016)
青稞(HordeumvulgareL. var.nudumHook. f.)为禾本科大麦属植物,属于栽培大麦的变种,因其成熟时籽粒内外稃与颖果分离,籽粒裸露,故称裸大麦[1]。青稞以其独特的营养结构和保健作用而成为极具开发利用价值的高原特色农作物之一[2],主要种植于我国西北和西南高原地区(西藏、青海、甘肃、四川、云南等)。种植区域内高寒缺氧,环境恶劣,降雨量少、蒸发量大[3],但青稞在如此恶劣的条件下表现出较强的适应性,说明其基因组中蕴藏了丰富的与抗旱和抗寒相关的优良基因。开发利用这些基因,不仅对选育青稞抗逆新品种具有重要理论意义,而且对作物抗胁迫基础研究具有促进作用。
干旱严重影响农作物的生长发育和产量,尤其是在高原地区[4-5],因而与抗旱相关的基因及相关蛋白成为研究的热点。HVA1基因与LEA蛋白、blt基因与LTP蛋白为其中的两大类。有研究表明,LEA 蛋白与细胞耐逆性有关[6],特别是与失水抗性有紧密联系[7]。根据蛋白中氨基酸序列的同源性及一些特殊基元序列(TAQAAKEKAGE),可将LEA蛋白分为6组[8]。而HVA1基因编码的LEA3蛋白,在植物遇到干旱、盐、低温和外源ABA胁迫时会大量表达,从而抵抗逆境胁迫[9-10]。另有研究表明,LTP 蛋白在抵御非生物胁迫过程中也发挥着重要的作用,LTP基因在受低温、干旱、外源ABA以及损伤等逆境胁迫时也会过量表达[11]。在大麦中blt(low-temperature responsive barley gene) 基因家族是一类低温反应基因,受低温诱导表达[12],其中blt4也受干旱和ABA的诱导[13]。
本实验室前期克隆了青稞HVA1和blt4.9基因,并发现在模拟干旱胁迫下这两个基因在抗旱性强的青稞品种中表达量显著高于抗旱性弱的品种[13-14],推测两基因均与青稞的抗旱性有关,但并没有转基因进行功能验证,也没有比较二者在青稞抗旱过程中作用的强弱,更不了解二者的抗旱反应机制。本研究以青稞为材料,从8个麦类作物常用候选内参基因 (DHN1、GAPDH、Actin-1、Actin-2、18SrRNA-1、18SrRNA-2、TC139057和PKABA) 中筛选到稳定表达的内参基因,并研究青稞HVA1和blt4.9基因对模拟干旱胁迫的响应,比较他们对抗旱性响应的差异,并在转基因拟南芥中验证了其抗旱性,初步探讨了二者在抗旱育种中的应用,以期为进一步开展青稞抗旱的分子机制研究奠定了基础。
1 材料与方法
1.1 试验材料
供试青稞品种为昆仑17号、昆仑12号和北青1号,由青海大学林科学院作物所青稞研究室繁育保存,三个品种的抗旱性强弱顺序为昆仑17号>昆仑12号>北青1号。
1.2 处理方法
精选籽粒饱满的青稞种子于清水中浸泡 1 h,0.5%高锰酸钾溶液消毒30 min,清水冲洗干净后浸入蒸馏水中,于4 ℃冰箱放置24 h。之后,各取20 粒种子播种于铺有 3 层蒸馏水浸润滤纸、直径为10 cm的培养皿内,置于人工气候箱中培养,温度为(25±1) ℃,光照度为433 mol·m-2·s-1,每天光照14 h,其间每天向滤纸加数滴蒸馏水 (约5 mL),使滤纸浸透并稍有剩余,以避免干旱。待幼苗长至约10 cm高时(约2周) 对昆仑12号幼苗进行15%PEG胁迫,分别于1、24、48、96、120和144 h取叶片提取RNA,同时采用不同浓度的PEG (1%、5%、10%、15%、20%、25%和30%) 和ABA (1、5、10、50、100、200和500 mol·L-1)胁迫;另外对昆仑17号、昆仑12号和北青1号幼苗进行PEG (1%、5%、15%和25%) 胁迫,24 h后分别取叶片提取RNA,并测定生理指标。
1.3 引物设计
利用Primer Primier5.0软件,根据文献中常用的内参基因序列设计引物1~8;根据实验室已克隆的青稞HVA1和blt4.9基因的序列,在开放阅读框 (ORF) 内设计荧光定量PCR引物9和10 (表1)。引物由生工生物工程 (上海) 有限公司合成。
1.4 Real-time PCR
采用iQ5荧光定量PCR仪(Bio-Rad,美国),按照SYBR Premix Ex Taq Ⅱ (Takara,日本) 说明书分别建立目的基因和内参基因的反应体系。其组成:cDNA 50.0 ng,10 μm·L-1,上下游引物各0.5μL,SYBR○RPremix Ex TaqTMⅡ (TliRNaseH Plus) 12.5 μL,加ddH2O至25 μL。反应参数为95 ℃ 3 min; 95 ℃ 10 s, 61 ℃ 30 s, 95 ℃ 1 min, 61 ℃ 1 min, 40 个循环。
1.5 转青稞 HVA1和 blt4.9拟南芥植株的获得
利用本实验室构建的HVA1和blt4.9植物表达载体质粒转化农杆菌GV3101菌株,挑单菌落培养进行菌液PCR鉴定,获得阳性菌株。然后选取健壮的拟南芥植株,以花序法转化Columbia 生态型拟南芥。侵染植株收获T0代种子后,铺洒在25 mg·L-1的潮霉素抗性MS固体培养基上,4 ℃暗培养 2 d后 22 ℃、16 h 光照培养。将正常生长的抗性苗移栽到土壤中,成活后提取DNA,利用引物11和12 (表1)进行目的基因PCR扩增检测,保留阳性植株。再经过上述方法连续筛选得到阳性苗(T2代)用于后续试验。
1.6 基因表达分析
采用2-△△Ct法计算基因相对表达量[15], 1%~30% PEG模拟干旱胁迫,以HVA1基因在1% PEG胁迫处理下的表达量为1;15% PEG模拟干旱胁迫1~144 h,以HVA1基因在胁迫处理1 h的表达量为1;ABA胁迫处理,以HVA1基因在1 μmol L-1ABA处理下的表达量为1;HVA1、blt4.9基因在不同抗旱性品种中的差异,以昆仑17号HVA1和blt4.9基因在1%PEG胁迫处理下的表达量为1。
1.7 生理指标测定
蛋白质含量以考马斯亮蓝G250染色法[16]测定;相对电导率采用电导率仪法测定,相对电导率=(浸泡电导率值/煮沸后电导率值)×100%[17];丙二醛 (MD)含量采用硫代巴比妥酸显色法测定[18]。
1.8 数据处理
用Stst 2与SPSS 13.0进行方差分析和多重比较。用Excel作图。
2 结果与分析
2.1 模拟水分胁迫下青稞内参基因的筛选结果
由于模拟干旱胁迫下青稞叶片中能够稳定表达的内参基因未见报道,为保证研究结果的准确性,本研究首先进行了模拟干旱胁迫下内参基因的筛选。由图1可知,以昆仑12号叶片总RNA反转录的cDNA为模板,用RT-PCR扩增各内参基因片段,结果,15%PEG模拟干旱处理1、24、48、96、120和144 h后各内参基因的表达产生了明显的不同,1~5和8号内参基因在48~120 h内偶有表达,1~24 h和144 h均不表达;而6和7号内参基因在1~144 h内均大量表达,且与引物设计的预期片段大小一致,说明这两个基因扩增反应具有较高的专一性。Gel-PRO analyzer分析结果表明7号内参基因较6号内参基因表达量变化幅度小,故用7号内参基因检测模拟水分胁迫下青稞HVA1和blt4.9基因的荧光定量PCR 分析。

表1 所用引物名称及序列Table 1 Primers used to determine the expression of different genes using real-time PCR in this study
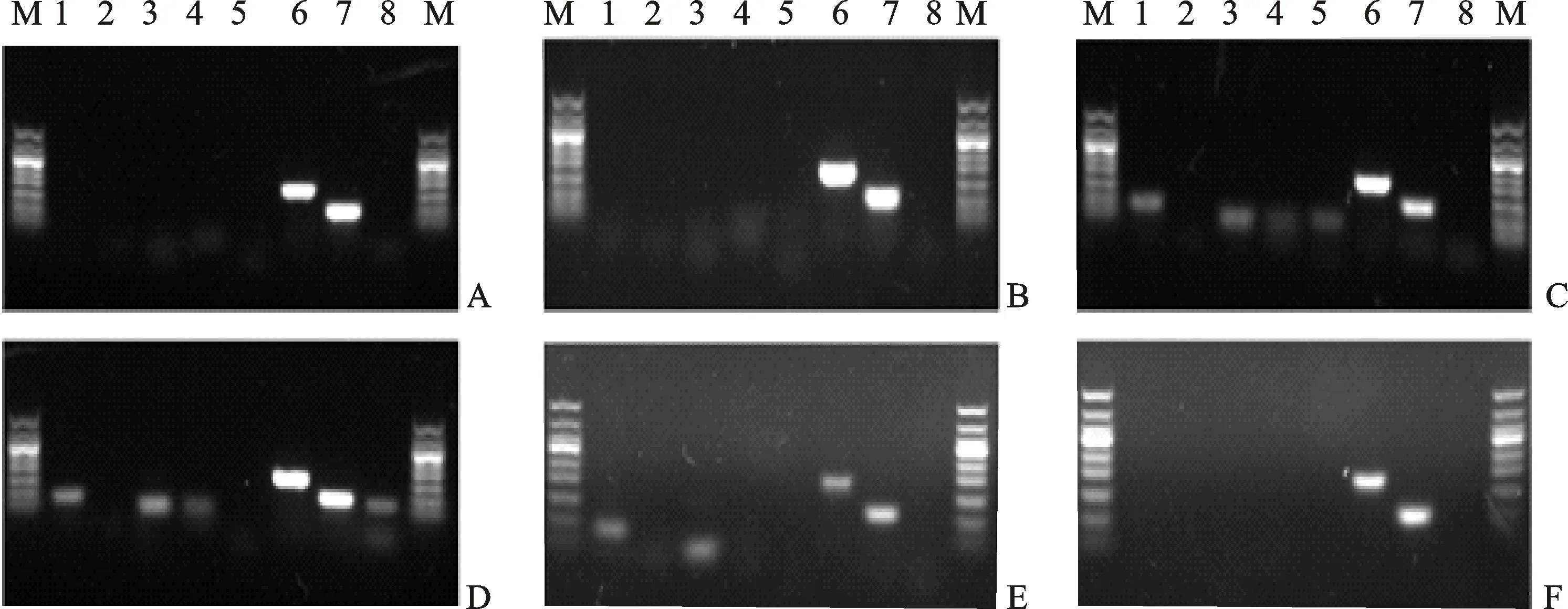
A~F分别为15%PEG模拟干旱处理1、24、48、96、120和144 h后各内参基因的PCR扩增结果;1~8分别为内参基因DHN1、GAPDH、Actin-1、Actin-2、18SrRNA-1、18SrRNA-2、TC139057、PKABA的扩增结果;M为Marker。
A, B, C, D, E and F stand for the amplification result of internal reference genes under 15% PEG drought stress for 1, 24, 48, 96, 120 and 144 h, respectively.1, 2, 3, 4, 5, 6, 7 and 8 stand for the amplification result of housekeeping genesDHN1,GAPDH,Actin-1,Actin-2,18SrRNA-1,18SrRNA-2,TC139057andKABA,respectively.M:Marker.
图1 15%PEG模拟干旱胁迫下内参基因的表达
Fig.1 Expression of housekeeping genes under 15% PEG drought stress
2.2 青稞 HVA1、 blt4.9基因对模拟干旱胁迫的响应
以TC139057为内参基因,检测了青稞HVA1和blt4.9基因在不同浓度PEG处理及15%PEG模拟水分胁迫1~144 h的表达量。结果(图2)表明,在不同浓度PEG模拟水分胁迫下,随着胁迫浓度的提高,青稞两基因表达量先提高后降低,其中25% PEG模拟水分胁迫下的基因表达量最高,与对照(1%PEG处理,下同)差异极显著 (P<0.01),且HVA1基因在10%~20% PEG胁迫下表达量显著(P<0.05)高于blt4.9基因,而在25%~30%PEG胁迫下显著低于blt4.9基因 (P< 0.05);在15%PEG模拟水分胁迫下,随着胁迫时间的延长,两基因的表达量先提高后降低,其中HVA1基因在胁迫48 h时表达量最高,blt4.9基因在胁迫96 h时表达量最高且与对照差异极显著 (P<0.01), 96 h后HVA1基因表达量显著低于blt4.9基因 (P<0.01)。可见,青稞HVA1和blt4.9基因对PEG模拟水分胁迫均有不同程度的响应,但响应方式不同,较低浓度胁迫和胁迫处理时间较短时HVA1基因表达量高于blt4.9基因,反之低于blt4.9基因。
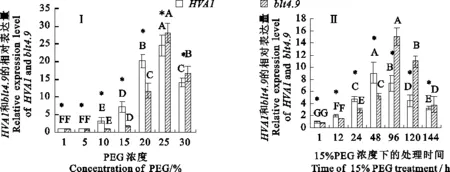
图柱上的大写字母不同表示同一基因在不同浓度PEG处理之间(图Ⅰ)或者在不同处理时间之间(图Ⅱ)有显著性差异(P<0.01),*号表示同一处理下blt4.9基因与HVA1基因间有显著差异(P<0.05),下同;分别以1%PEG处理和1 h处理下HVA1基因和blt4.9基因表达量为对照。图3同。
Capital letters above the column indicate significant differences under different treatments at 0.01 level. * indicates significantly different among the expression levels ofblt4.9andHVA1in a same treatment at 0.05 level.The expression levels ofHVA1andblt4.9genes treated with 1% PEG or 1 h were taken as controls. The same in figure 3.
图2 青稞HVA1、blt4.9基因在不同 PEG浓度处理下(图Ⅰ)以及15%PEG处理0~144 h(图Ⅱ)的表达量
Fig.2Expression levels ofHVA1andblt4.9genes under different PEG concentrations(Fig.Ⅰ) and under different time treated with 15% PEG(Fig.Ⅱ)
2.3 青稞 HVA1、 blt4.9基因对ABA的响应
有研究表明,干旱胁迫往往引起ABA含量的变化[19]。本实验测定了1~500 μmol·L-1ABA处理下青稞HVA1和blt4.9基因表达量的变化。结果(图3)表明,随着外源ABA浓度的增加,两基因的表达量均先增加后降低,其中HVA1基因在10 μmol·L-1ABA处理下的表达量最高,为对照(1 μmol·L-1ABA处理,下同)的7.72倍,且与对照的差异极显著(P< 0.01);blt4.9基因在50 μmol·L-1ABA处理下表达量最高,为对照的8.96倍,且与对照的差异极显著(P<0.01);在较低浓度的ABA(1~10 μmol·L-1)处理下,HVA1基因表达量显著高于blt4.9基因,反之低于blt4.9基因(P<0.05)。可见,青稞HVA1和blt4.9基因对不同浓度的ABA处理会产生不同程度的响应。
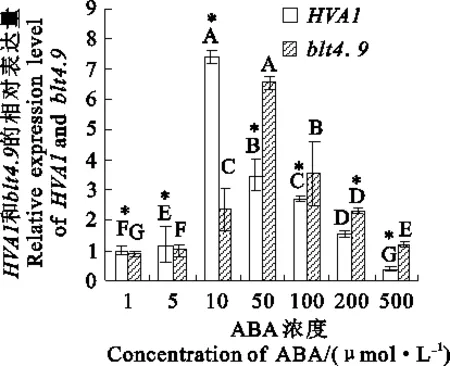
以1 μmol·L-1ABA处理下HVA1和blt4.9基因表达量为对照。
The expression levels ofHVA1andblt4.9genes under 1 μmol·L-1ABA treatment were used as controls.
图3青稞HVA1和blt4.9基因在不同浓度ABA处理下的表达量
Fig.3Expression level ofHVA1andblt4.9genes under ABA stress
2.4 转青稞 HVA1、 blt4.9基因的拟南芥抗性筛选及分子检测结果
在25 mg·L-1的潮霉素抗性培养基上,转化成功的拟南芥正常生长,而未转化成功的拟南芥逐渐白化死亡。将抗性培养基筛选得到的T2代幼苗移栽到培养钵中继续培养成苗,剪取拟南芥叶片提取基因组DNA,用目的片段引物进行PCR检测,1%琼脂糖电泳后出现目的条带(图4),表明HVA1和blt4.9基因已经成功整合到拟南芥基因组中。筛选到的阳性植株可以用于抗旱性鉴定实验。
2.5 转青稞 HVA1、 blt4.9基因的拟南芥抗旱性鉴定结果
以转青稞HVA1、blt4.9基因的拟南芥株系 (HVA1、blt4.9) 和对照植株 (野生型WT) 的离体叶片为材料,测定了0~20%PEG模拟干旱胁迫处理下可溶性蛋白质含量、电解质渗透率和丙二醛(MDA)含量,结果如表2所示。随着PEG浓度的升高,转基因植株和对照植株的叶片可溶性蛋白质含量都显著下降,而电导率和丙二醛含量都显著升高 。在同一PEG浓度处理下,转基因株系的叶片可溶性蛋白质含量与对照相差无几,而相对电导率和丙二醛含量均显著低于对照。可见两基因转入拟南芥中均不同程度的提高了拟南芥的抗旱性。
2.6 青稞 HVA1、 blt4.9基因表达在不同抗旱性品种中的差异
以抗旱性不同的青稞品种为材料,检测了其在15%和25%PEG模拟水分胁迫处理下青稞HVA1、blt4.9基因的表达量 (表3)。结果表明,两种模拟水分胁迫处理下青稞HVA1、blt4.9基因的表达量均显著高于对照(P<0.01),且三个青稞品种之间两基因表达量的差异均达显著水平(P<0.01),高低顺序均为昆仑17号>昆仑12号>北青1号,与其实际抗旱能力相一致。另外,昆仑17号、昆仑12号和北青1号在15%PEG模拟水分胁迫下HVA1基因表达量的比例为 2.88∶1.76∶1,在25%PEG模拟水分胁迫下HVA1基因表达量的比例为1.63∶1.32∶1;而blt4.9基因表达量的比例分别为1.82∶1.30∶1和 1.95∶1.48∶1。可见,利用青稞HVA1基因在较弱水分胁迫下的表达量高低判断青稞的抗旱性强弱,比用blt4.9基因的效果好,而在较强水分胁迫下用blt4.9基因效果更佳。

H1~H10和B1~B4:转基因拟南芥;WT:野生型拟南芥;NC:提取空白对照;-:PCR空白对照;+:阳性质粒;M:marker。
H1-H10 and B1-B4:transgenicArabidopsis; WT:wildArabidopsis;NC:blank control of extraction; -:blank control of PCR;+:positive plasmids; M:marker.
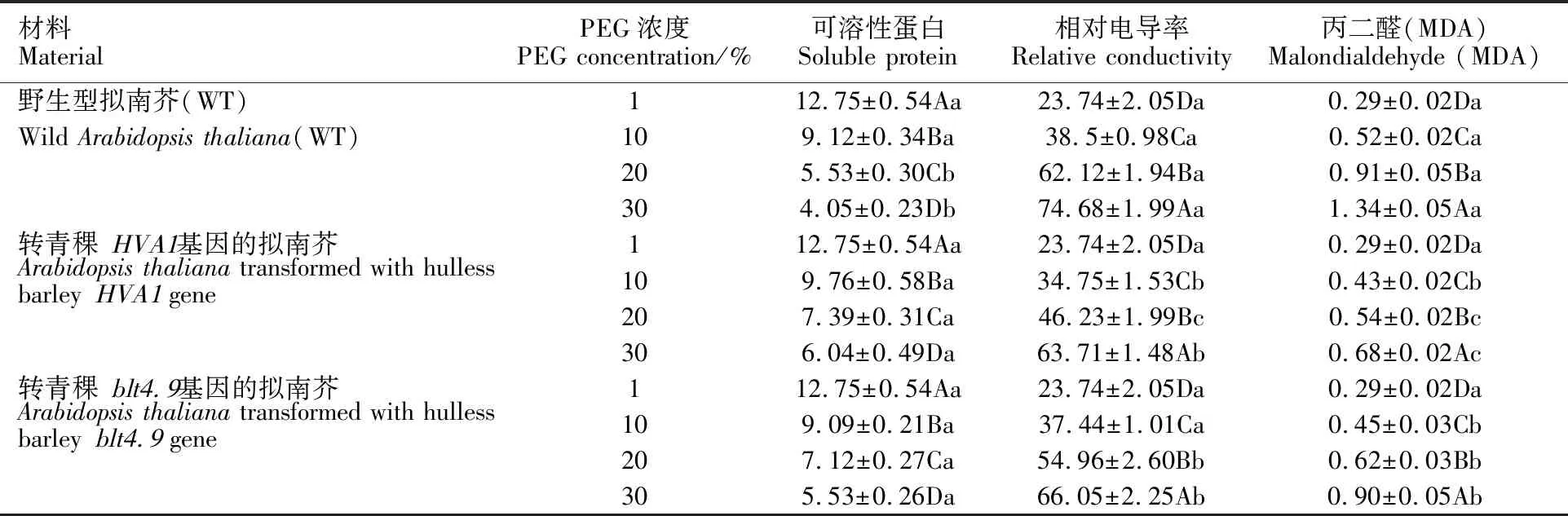
图4 转基因拟南芥的PCR检测结果
同一列数据后的不同大写字母表示同一实验材料在不同浓度PEG处理之间有显著性差异(P<0.01);同一列数据后的不同小写字母表示同一PEG浓度下不同实验材料之间有显著性差异(P<0.05)。
Different capital letters following data indicate significant differences among concentrations of PEG in the same experiment material at 0.01 level; Different lowercase letters indicate significant differences among experiment materials in the same concentrations of PEG at 0.05 level.
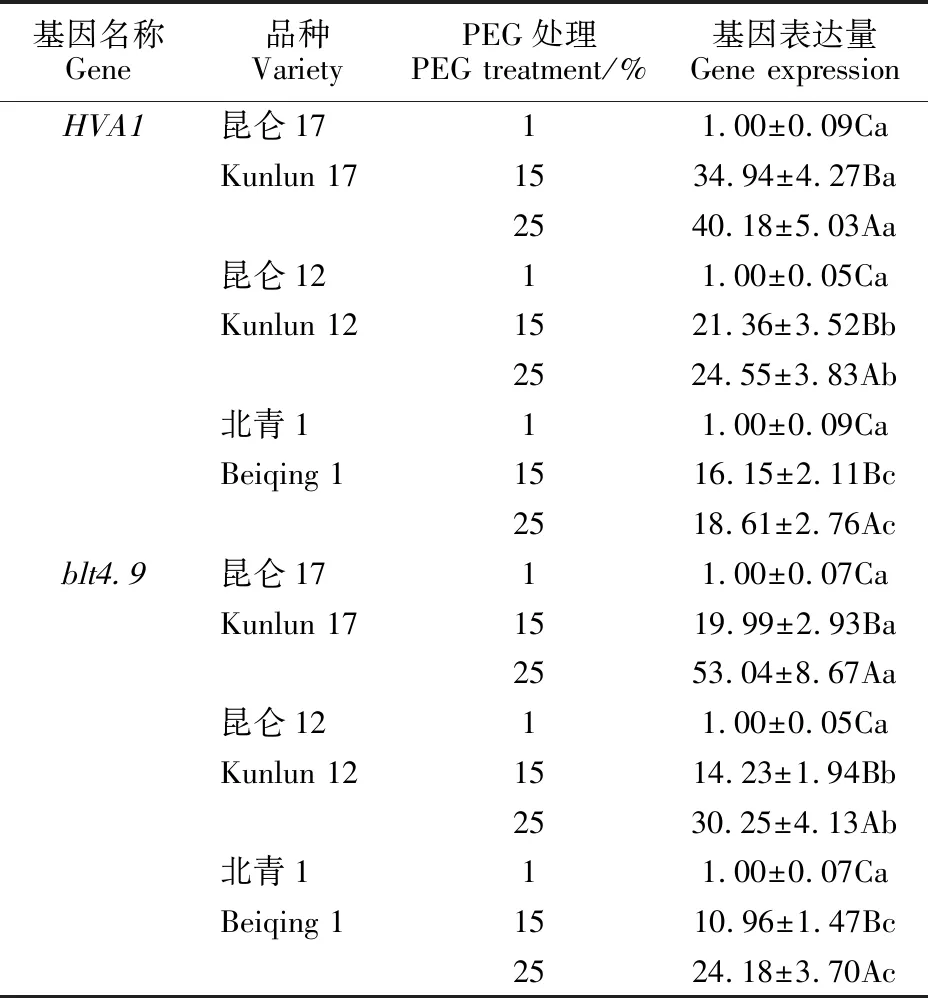
表3 模拟水分胁迫处理下抗旱性不同的青稞 HVA1、 blt4.9基因表达量Table 3 Expression of HVA1 and blt4.9 genes in huless barleys with different drough resistance under simulated drought stress
数据后的不同大写字母表示同一品种同一基因的表达量在不同浓度PEG处理之间有显著性差异(P<0.01);数据后的不同小写字母表示同一PEG浓度下同一基因在不同品种之间的表达量有显著性差异(P<0.01)。
Different capital letters following data indicate significant differences among concentrations of PEG in the same variety and the same gene at 0.01 level; Different lowercase letters indicate significant differences among varieties in the same concentrations of PEG and the same gene at 0.01 level.
3 讨 论
干旱胁迫是作物面临的最为常见的非生物胁迫类型,随着分子生物学技术的发展,干旱基因表达的检测是干旱胁迫研究的一个重要方面[20]。实时定量PCR技术是检测逆境下基因表达量的一个有效手段,在进行实时定量PCR 检测中,由于样品的RNA质量、反转录时cDNA合成的效率以及PCR的反应条件等都有可能存在差异,因此必须引入拷贝数高、稳定表达的内参基因对目标基因的表达量进行校正[21]。目前在大麦中已经有了胁迫条件下内参基因筛选的报道,在干旱和盐胁迫中大麦实时定量PCR内参基因的筛选研究结果表明,在进行实时定量PCR 研究之前需根据具体的实验条件先对内参基因进行筛选[22]。而且近年来,很多报道表明一些常用的内参基因在不同的组织、器官、发育时期或不同的实验条件下表达并不稳定[23-24]。本实验对15%PEG模拟干旱胁迫1~144 h下青稞叶片中8个内参基因表达的稳定性进行了检测,也得到了类似的结果,研究发现胁迫时间较短(1~24 h)和较长(144 h)时,除6号内参基因和7号内参基因外,大部分内参基因表达不稳定,可见,在进行实时定量PCR 研究时,筛选表达稳定、适宜特定实验条件的内参基因是获得准确可靠实验结果的重要前提[21]。
植株生物量和含水量是评价作物抗旱性的重要指标,作物的抗旱性具有复杂的机制,不能简单通过单一性状或者生理指标基因来评价青稞的抗旱性。近年来的研究发现了大量与抗旱相关的基因[25-27],以HVA1基因和大麦blt基因家族各成员为例,Xu等[25]、Sivamani等[26]和Lai等[27]通过转基因分析表明,HVA1基因过表达后可显著提高转基因水稻、小麦和桑树的抗旱及耐盐能力,这些研究为支持HVA1基因在植物抗水分和盐胁迫时起保护作用的假说提供了直接的证据;近年来研究表明,大麦blt基因家族是一类低温反应基因,但个别基因(如blt4)也同时受干旱和ABA 的诱导[30]。本实验室克隆了HVA1和blt4.9基因,并初步证明它们与青稞的抗旱性有关[13-14]。本实验比较了HVA1和blt4.9基因在模拟干旱胁迫下的表达量,结果表明,两基因的表达量呈先增加后下降的趋势,其原因可能是在受到轻度干旱胁迫或干旱时间较短时,植物体需要较多的LEA蛋白和LTP蛋白来稳定和修复膜系统,以便复水时可以迅速恢复细胞膜系统和正常的代谢能力;而在干旱较严重或者胁迫时间较长时,植物整个代谢系统因受到严重的水分胁迫而制约了两个基因的表达。是否如此,还需要通过测定其编码的LEA蛋白和LTP蛋白的含量来进一步验证。另外,受到轻度干旱胁迫或干旱胁迫时间较短时,HVA1基因表达量显著高于blt4.9基因;而受到重度干旱胁迫或干旱胁迫时间较长时,HVA1基因表达量显著低于blt4.9基因。同样在转基因拟南芥中也发现了此规律,两基因转入到拟南芥后测定叶片的可溶性蛋白质含量、电解质渗透率和丙二醛(MDA)含量值发现,在较低浓度(15%)PEG胁迫下三个抗旱相关指标在转HVA1拟南芥中优于转blt4.9基因材料;而在较高浓度(25%)PEG胁迫下则相反。可见HVA1基因对干旱胁迫较敏感,在干旱胁迫早期或轻度干旱即开始响应;而blt4.9基因对干旱胁迫较迟钝,在干旱胁迫后期或严重干旱时才开始响应,原因可能是两个基因或其编码的蛋白对干旱胁迫的响应机制不同,具体机制需要进一步验证。
另外, 本研究还发现, 在相同的逆境胁迫下抗逆性强的青稞品种,其HVA1和blt4.9基因表达量显著高于抗逆性弱的品种,而且较低浓度(15%)PEG胁迫下三个品种HVA1基因表达量的差异比较高浓度(25%)PEG胁迫下的大,而blt4.9基因则正好相反。这提示我们在受到较轻干旱胁迫时可以通过检测HVA1基因的表达量来比较青稞的抗旱性;在受到较重干旱胁迫时可以通过检测blt4.9基因的表达量来比较青稞的抗旱性。
