循环水养殖模式下鱼生长对水环境因子的响应模型构建
2019-07-23程香菊具家琪胡佳纯余德光
程香菊,具家琪,胡佳纯,谢 骏,余德光
循环水养殖模式下鱼生长对水环境因子的响应模型构建
程香菊1,具家琪1,胡佳纯2,谢 骏3,余德光3
(1. 华南理工大学土木与交通学院,广州 510641; 2. 广东省水利水电科学研究院,广州 510610;3. 中国水产科学研究院珠江水产研究所,广州 510380)
为探究鱼类生长对水环境的响应,预测鱼类在养殖水环境多因子协同作用下的生长速度,进行了室内曝气推流循环水养殖罗非鱼试验,试验持续周期为8周。结果表明,在一定范围内,随着溶解氧质量浓度的增加,鱼的食物转化效率和特定生长率均有所提高;随着非离子氨质量浓度的增加,鱼的食物转化效率和特定生长率均有所降低;而亚硝酸盐质量浓度由于变化不大且均处于安全质量浓度范围,该试验中对鱼的食物转化效率和特定生长率未产生显著影响。基于这一系列试验结果对罗非鱼特定生长率进行了非线性拟合,建立了鱼的生长预测模型,2为0.82,并通过实测数据验证了模型的有效性和普适性。预测模型表明,养殖初始鱼质量、养殖密度、非离子氨以及亚硝酸盐质量浓度的增加,均会导致鱼生长速度减缓,而提高溶解氧质量浓度则可以提高鱼生长速度。该预测模型虽然是在曝气推流循环养殖模式下获得的,但对其他养殖模式同样适用,使鱼生长对水环境因子的响应变得可测,为促进养殖鱼类的健康发展、养殖系统的优化和养殖效益的提高提供了便利和参考。
水产养殖;鱼;生长;水环境;预测模型;非线性拟合
0 引 言
随着养殖规模的提高,传统水产养殖模式的弊端不断显现,包括对水资源的浪费、污染等。池塘曝气推流循环养殖系统则是在此基础上对高效节能水产养殖模式的探索,在国外得到了大力推广和快速发展[1-2],且自2013年从美国引入中国以来,已发展到全国的18个省市[3]。该养殖模式相较于传统养殖模式,具有使养殖水体循环流动、补充溶解氧、提高鱼类成活率,同时降低水资源消耗和减少污染等优点[4]。
大量研究表明,溶解氧、氨氮和亚硝酸盐是影响鱼类生长的三大影响因子。溶解氧通过限制代谢率进而影响鱼类的生长[5],是水质状况的重要指标。Tranduy等[6]研究发现,罗非鱼在3.0 mg/L的低氧环境中摄食量下降、生长缓慢。研究表明黄颡鱼[7]、大西洋比目鱼[8]在低氧环境下也会生长减缓。氨氮对水生动物有一定的毒害作用,质量浓度过高会对鱼体内酶的催化作用和细胞膜的稳定性产生严重影响,并破坏排泄系统和渗透平衡[9]。Paust等[10]报道较高的非离子氨质量浓度(≥0.17 mg/L)会导致大比目鱼食欲下降、生长速度减缓和血液生化指标发生明显变化。类似非离子氨对生长的抑制作用在斑点狼鱼[11]和罗非鱼[12]均被证实。过高质量浓度的亚硝酸盐会导致鱼虾血液中的亚铁血红蛋白被氧化成高铁血红蛋白,而后者不能运载氧气,从而抑制血液的载氧能力,造成组织缺氧,鱼类摄食能力低甚至死亡[13]。类似亚硝酸盐对生长的抑制作用在大西洋鳕鱼[14]和虹鳟鱼[15]均被证实。溶解氧、氨氮和亚硝酸盐三者间还会产生协同作用,溶解氧不足,非离子氨和亚硝酸盐质量浓度高,会使鱼类机体代谢功能失调,免疫力下降,导致鱼病发生[16]。
针对循环水养殖系统下水环境对鱼类的生长影响,目前国内外对水环境单因子条件的研究较多,而对溶解氧、非离子氨和亚硝酸盐协同作用下生长模型的研究少有报道。因此,充分分析并预测鱼生长对水环境因子的响应,对养殖产量的提高是非常必要的。
鉴于此,本文根据野外池塘曝气推流循环养殖系统的原型,按照一定的几何比尺,自行设计了一套室内曝气推流循环水养殖模型,以罗非鱼为养殖对象,在养殖水槽内圈养8周,通过改变曝气流量,观察养殖期间罗非鱼的生长情况,定期称量鱼的质量以及测量溶解氧、非离子氨、亚硝酸盐等水质参数,探讨水环境多因子协同作用对鱼生长的影响,并构建鱼生长预测模型,以期为促进养殖鱼类的健康发展、养殖系统的优化和养殖效益的提高提供参考。
1 试验装置与方法
1.1 室内循环水养殖装置
室内循环水养殖模型及设计尺寸如图1所示,其中,图1a为养殖循环水槽俯视图,图1b为养殖循环水槽侧视图,主要包括:曝气推流装置①、养殖水槽单元②、清水区域③、走道④。整个模型装置尺寸为370 cm×310 cm×75 cm(长×宽×高)。

① 气推流装置 ②养殖水槽单元 ③清水区域 ④走道
①Aeration plug-flow device ②Aquaculture tank unit ③Clear water area ④Aisle
注:为底坡坡度。
Notes:is the slope of the bottom slope.
图1 室内池塘循环水养殖模型示意图
Fig.1 Schematic diagram of indoor pond circulating water aquaculture model
曝气推流装置的设计如图2所示,主要包括支架1、曝气装置2和弧形挡板3。支架1用于支撑并连接弧形挡板和曝气装置。支架的长宽高为正方形,边长30 cm。同时,曝气装置2安装在支架的正下方,主要由一组输气管道和5根微气泡发生管组成,微气泡发生管的物理性质如文献[17]所述。养殖水槽单元②详细结构和尺寸如图3所示,底坡坡度=0.01。把该装置固定卡放在养殖水槽入口,开启空气压缩机,调节压力和气体流量大小,持续产生大量上浮微气泡,微气泡被上方的弧形挡板所阻挡,迫使微气泡改变原来上浮的路径,从弧形挡板的前方流出,从而带动水体自下而上、自水槽入口到出口的流动。

图2 曝气推流装置示意图
1.2 试验鱼及饲养管理
试验鱼为同一品种同龄的尼罗罗非鱼,平均规格为:体长(8.93±0.34) cm、质量(23.61±3.50) g。为了使罗非鱼适应试验室的养殖环境,在试验开展前将健康无疾病、活力强、规格整齐的罗非鱼暂养在500 L圆形塑料桶中7 d。采用已充分曝气的自来水,以保证自来水中氯气基本上完全散逸。暂养期间采用气泵增氧,并每日保持稳定的生长环境。

1.支架 2.曝气装置 3.弧形挡板
1.Frame 2.Aeration device 3.Arc baffle
图3 养殖水槽单元示意图
Fig.3 Schematic diagram of aquaculture tank unit
暂养后选取健康活泼且规格均匀的罗非鱼随机分组放入养殖水槽内(220 cm×30 cm×40 cm)进行正式试验,每个养殖水槽内养殖15尾鱼。试验以7 d为1个测量周期,共8个测量周期(56 d)。试验用水为已充分曝气的自来水。试验期间水温由加热棒控制,保持在(26.7±1.2)℃[18],pH值保持在7.0~8.0之间,期间不换水,每周仅添加去氯自来水用于补充蒸发和水质测定损失的水分。每天在08:30和20:30投喂罗非鱼膨化配合饲料,饲料含有约31%的粗蛋白、0.8%的总磷。每天2次的投喂总量为鱼质量的3%左右,投喂半小时后捞出养殖区残饵,计算残饵量及实际摄食量。同时根据每个水槽试验鱼的总质量调整下一周的饵料投喂量。
1.3 试验条件及样本采集
试验设置3组不同曝气流量的试验组,分别为0、30、50 L/min,每组曝气流量下设3个平行养殖水槽单元。溶解氧采用24 h不间断开启曝气推流系统的方法控制。分别在第0、7、14、21、28、35、42、49、56 天,测定水环境指标以及鱼质量。溶解氧采用YSI Pro ODO溶氧仪测定;pH值采用便携式pH计测定;氨氮采用靛酚蓝分光光度法测定。非离子氨根据式(1)计算[19]。
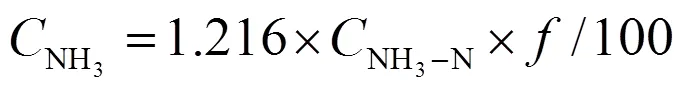

式中为水解常数,为酸度系数(=-lg()= 0.090 18+2 729.92/);pH值为实测值;为热力学温度(=1+273.15,1为试验水体温度,℃)。
用精度为0.01 g的电子称称量养殖水槽内所有鱼的质量,并取平均值为水槽鱼的质量。罗非鱼特定生长率和食物转化效率的计算公式如式(3)和(4)。
特定生长率(%/d)=(ln末−ln初)/2×100% (3)
食物转化效率(%)=(末−初)/×100% (4)
式中2为1个测量周期的天数,为7 d;末为测量周期的鱼质量,kg;初为该测量周期初的鱼质量,kg;为该测量周期鱼的总摄食量,kg。
2 试验结果与分析
2.1 水环境单因子对鱼生长的影响
图4a给出了鱼的特定生长率对溶解氧质量浓度的响应。从图4a可以看出,水中溶解氧质量浓度在1.3~7.0 mg/L范围内,随溶解氧质量浓度的增加,罗非鱼特定生长率增大。通过分析,溶解氧质量浓度与特定生长率之间的关系符合指数函数变化趋势,即:=0.231e0.384x,其决定系数2为0.62,表明相关性较好。
试验期间通过观察发现,在高溶解氧情况下,罗非鱼游动较快,摄食迅速,且摄食过程较为剧烈,常使水体剧烈扰动发出声音;而低溶解氧情况下,罗非鱼常游动缓慢,这与杨凯等[7]对黄颡鱼的研究结果一致:低氧会抑制鱼类的摄食活动。图4b给出了鱼的食物转化效率对溶解氧质量浓度的响应。从图4b可以看出,降低溶解氧质量浓度,会使食物转化效率下降,表明低食物转化效率是低溶解氧导致鱼类生长速度减缓的结果之一,与Abdel-Tawwab等[20]研究结果一致。
Thorarensen等[8]研究发现溶解氧质量浓度显著影响大比目鱼的生长速率,当氧饱和度从57%增加到100%时,鱼的生长速率逐渐增加,而氧饱和度的进一步增加不会改善生长速率。在本研究中,水体溶解氧质量浓度主要来源于曝气推流装置的微气泡界面与水体的氧传质以及自由表面的湍动传质,溶解氧质量浓度一般不会超过其饱和质量浓度。从测定结果看出,本研究的结论与Thorarensen等[8]的研究结果吻合,均表明在一定范围内溶解氧增加对鱼生长有利。

图4 鱼的特定生长率和食物转化效率对溶解氧质量浓度的响应
图5a给出了鱼的特定生长率对非离子氨质量浓度的响应。从图5a可以看出,水中非离子氨质量浓度在0.001 6~0.025 mg/L范围内,随非离子氨质量浓度增多,罗非鱼特定生长率减小。通过分析,非离子氨质量浓度与特定生长率之间的关系符合指数函数变化趋势,即:=3.42e-113.5x,其决定系数2为0.54,表明相关性较好。

图5 鱼的特定生长率和食物转化效率对非离子氨质量浓度的响应
水环境中氨氮质量浓度上升会抑制鱼体的氨氮排泄[21],同时非离子氨会通过鱼鳃进入鱼体,使血液和组织中氨氮的质量浓度升高,进而对鱼体产生一系列毒性影响[22]。氨氮对水生动物的毒性主要来源于非离子氨,远高于离子氨约300~400倍[23]。Foss等[11]研究用0.39 mg/L的非离子氨处理斑点狼鱼,发现斑点狼鱼的平均质量显著低于0.000 6和0.13 mg/L处理下的试验组鱼,且随非离子氨质量浓度的增加,摄食量和食物转化效率均下降。图5b给出了鱼的食物转化率对非离子氨质量浓度的响应。从图5b可以看出,非离子氨质量浓度增加,会使食物转化效率下降,与Foss等[11]研究结果一致。本研究结果表明,在养殖过程中,非离子氨增加导致了罗非鱼生长速度减缓,因此建议定期更换新鲜水体,以确保养殖产量。
图6a、图6b分别给出了鱼特定生长率和食物转化效率对亚硝酸盐质量浓度的响应。从图6a、6b可以看出,水中亚硝酸盐质量浓度在0.05~0.19 mg/L范围内,点比较分散,且并未呈现出明显的规律性,因此本试验中亚硝酸盐质量浓度对罗非鱼特定生长率和食物转化效率均未产生显著影响。然而,Siikavuopio等[14]研究发现,将大西洋鳕鱼养殖在亚硝酸盐质量浓度为0、1、2.5、5 mg/L的水环境中,31 d后生长速度随质量浓度升高而降低。本研究可能是由于各试验组亚硝酸盐质量浓度均低于0.2 mg/L,处于安全质量浓度,罗非鱼能够适应。
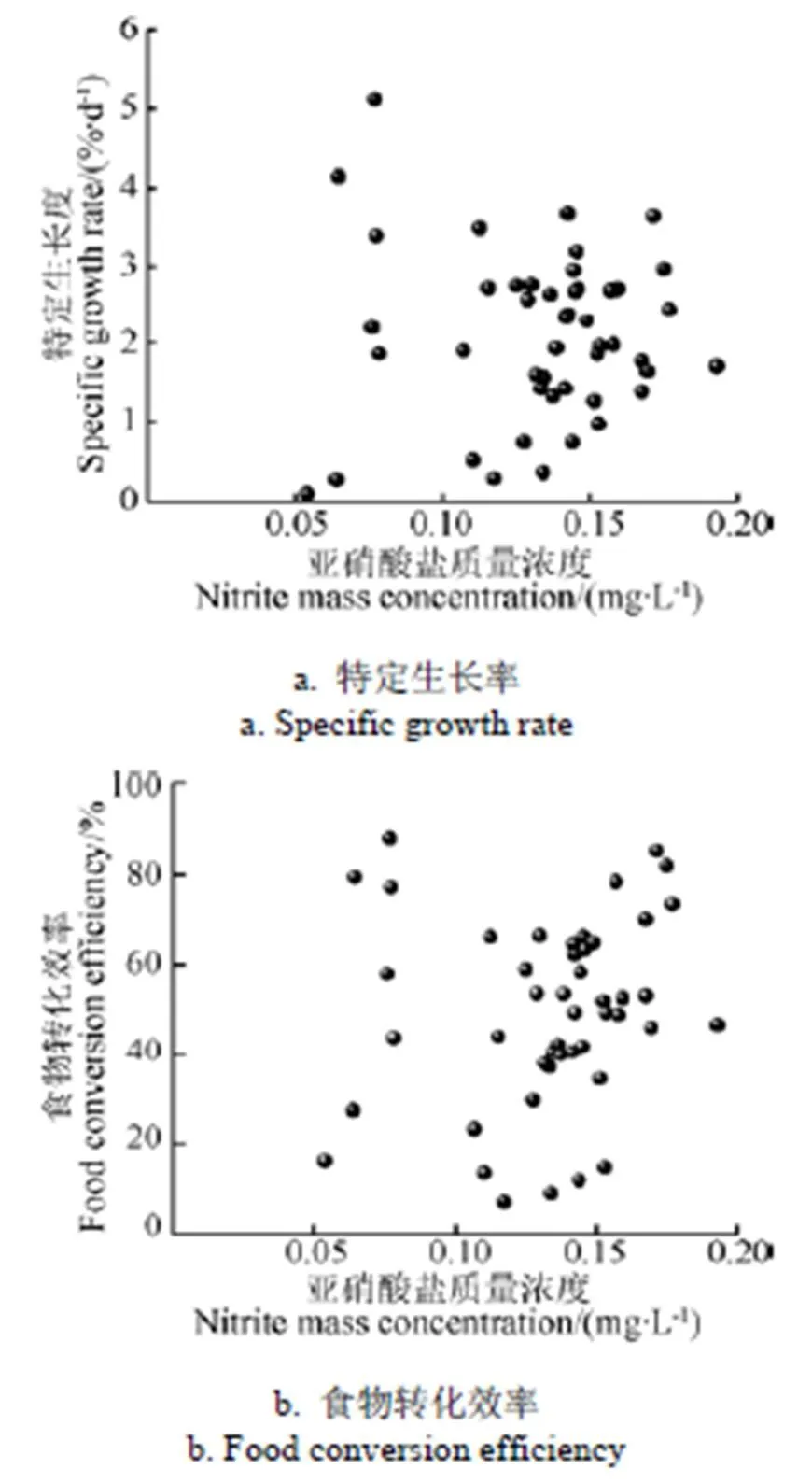
图6 鱼的特定生长率和食物转化效率对亚硝酸盐质量浓度的响应
2.2 水环境因子协同作用下的鱼生长预测模型
从2.1节可以发现,仅根据某个水环境单因子建立与鱼特定生长率的关系拟合程度不高,因此需要考虑各水环境因子的协同作用对鱼特定生长率的影响。同时养殖水体中水环境因子的变化具有高度非线性和不确定性,使得鱼生长与各环境因子之间并不呈现出简单的线性关系。因此,本研究随机取36组试验结果为样本建立模型,取另外12组试验结果验证模型。模型根据每一个养殖周期,选择每一个养殖周期初始罗非鱼质量、初始养殖密度以及养殖期间溶解氧、非离子氨和亚硝酸盐平均质量浓度作为响应因子,以罗非鱼的特定生长率作为响应值,进行非线性拟合分析,得到罗非鱼特定生长率与水环境因子的关系如式(5)所示

其中,

式中表示水环境因子的协同作用因子;SGR为罗非鱼特定生长率,%/d;为初始养殖密度,kg/m3;为初始质量,kg;DO为溶解氧质量浓度,mg/L;NH3为非离子氨质量浓度,mg/L;NO2为亚硝酸盐质量浓度,mg/L。
式(5)的决定系数2为0.82,表明相关性较好,并且拟合散点和验证散点基本上都落在或围绕在拟合线周围(如图7所示)。

图7 鱼特定生长率对水环境因子的协同响应
从式(5)-(6)可以看出,鱼的生长对水环境因子的响应包括了5个因子的协同作用,即:、、DO、NH3、NO2。随着养殖初始鱼质量的增加,特定生长率减小,且呈幂函数关系,这符合鱼类前期生长速度快,后期生长速度慢的特性[24]。García等[25]研究发现海鲷的特定生长率和质量之间存在幂函数关系SGR=aW,关系式表示鱼在幼小时增长速度更快,与本文分析规律一致。随着养殖密度的增加,特定生长率减小。这是由于养殖密度过高,会胁迫鱼群产生一系列的生理生化变化,种内空间和饵料竞争加剧,为了相互避让、调节生理功能而消耗更多的能量,因而影响鱼的生长,并降低鱼类的生长速度[26]。同时养殖密度的提高还会带来水质恶化进而影响鱼类的生长和生存。同时预测模型表明,在一定范围内随着溶解氧质量浓度的增加,特定生长率与其呈指数关系增加[27]。而随着非离子氨质量浓度和亚硝酸盐质量浓度的增加,特定生长率均与其呈指数关系减小。
2.3 鱼生长预测模型的普适性分析和应用
从式(5)-(6)可以看出,SGR与存在一个线性关系,即

式中的计算式等同于式(6),为系数。
为了验证预测模型的普适性并将其推广应用,选取了5篇文献的工况对式(6)-(7)进行验证。将5篇文献中的试验条件(见表1)分别代入式(6)-(7),得出相对应的系数值和,即求得生长预测模型,列于表1最后一列。同时将得到的各工况预测SGR值与实测SGR值相对比(如图8所示),可以看出散点几乎都能落在=的直线上,说明预测模型可靠。

表1 生长预测模型的应用
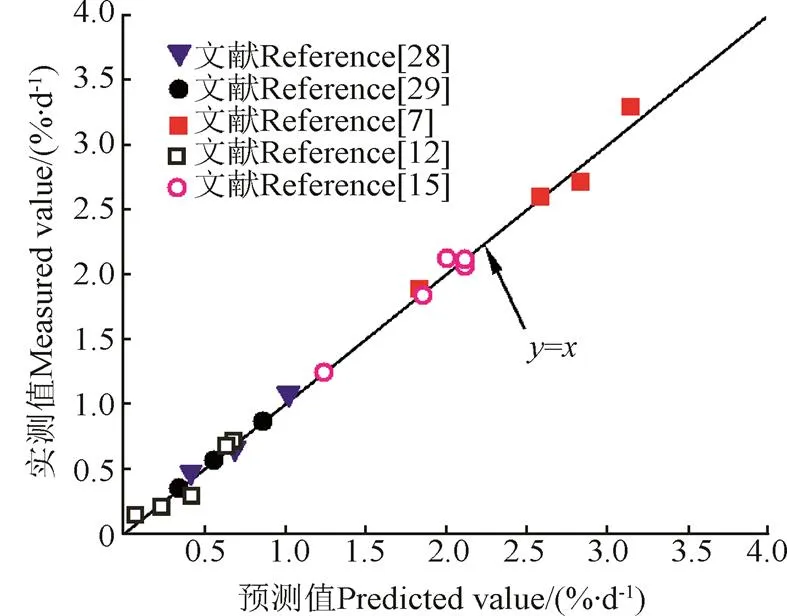
图8 鱼特定生长率(SGR)预测值和实测值的对比图
从以上分析可以发现,在实际养殖水域中,养殖鱼类的特定生长率与水域环境的定量响应可以由式(6)和式(7)确定。
本文生长预测模型的建立,确定了不同养殖密度、鱼初始质量、溶解氧、非离子氨和亚硝酸盐对鱼生长的影响程度,得出了不同水环境因子变化引起的鱼生长速度变化。在土地愈发紧张的现状下,为了提高养殖水平,集约化养殖多被采用,随着养殖密度的提高,将会引起水环境非离子氨和亚硝酸盐质量浓度增加,对鱼体产生生长速度降低等恶性影响,可以通过定期更换新鲜水体、补充溶解氧等措施来改善水体环境,保障鱼类健康的生长。在实际养殖生产中,本模型为通过及时调整养殖水域水环境相关因子,为快速找到最佳生长率奠定理论基础,有助于提高养殖效益。此外,水温也是影响鱼类健康生长发育的重要因子。不同鱼类有其适应的水温范围,使其摄食和生长指标均能达到较优值[30]。而本文生长预测模型并未考虑水温的影响,具有一定的局限性,还需进一步研究。
3 结 论
针对曝气推流循环养殖模式下罗非鱼的生长情况,进行了8周的养殖试验,计算了罗非鱼食物转化效率以及特定生长率指标,分析了鱼特定生长率对水环境因子的响应,并得到其生长预测模型。相关结论如下:
1)溶解氧质量浓度的增加能提高罗非鱼的食物转化效率和生长速度。因此在循环水养殖中,可以通过在一定范围内增加溶解氧的质量浓度来加快养殖速度。
2)非离子氨质量浓度的增加会导致罗非鱼食物转化效率和生长速度的降低。因此建议定期更换新鲜水体,降低水体中非离子氨的质量浓度,以确保养殖产量。
3)建立了养殖水体罗非鱼生长对水环境响应模型,2为0.82。模型表示在实际养殖过程中,可以基于养殖水环境因子使实际鱼的生长速度变得可测,可依据该预测模型及时调整水环境因子,使养殖鱼类的健康发展、养殖系统和养殖效益得到尽可能的最优化。
[1] Martins C I, Eding E H, Verdegem M C J, et al. New developments in recirculating aquaculture systems in Europe: A perspective on environmental sustainability[J]. Aquacultural Engineering, 2010, 43(3): 83-93.
[2] 贾丽,潘勇,刘帅. 池塘内循环流水养殖模式:美国的一种新型养殖模式[J]. 中国水产,2011(1):40-42.
[3] 周恩华. IPA池塘循环流水养鱼系统的心脏是增氧和推水[Z]. 中国水产养殖网,2017.
[4] 马立鸣,赵睿,范毛毛,等. 新型池塘循环流水养殖模式初探[J]. 中国水产,2016(1):78-83.
[5] Dupont-Prinet Aurélie, Vagner M, Chabot D, et al. Impact of hypoxia on the metabolism of Greenland halibut ()[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2013, 70(3): 461-469.
[6] Tranduy A, Schrama J W, Dam A A V, et al. Effects of oxygen concentration and body weight on maximum feed intake, growth and hematological parameters of Nile tilapia,[J]. Aquaculture, 2008, 275(1/2/3/4): 152-162.
[7] 杨凯,樊启学,张磊,等. 溶氧水平对黄颡鱼稚鱼摄食、生长及呼吸代谢的影响[J]. 淡水渔业,2010,40(2):24-29.
Yang Kai, Fan Qixue, Zhang Lei, et al. Effects of dissolved oxygen on feed intake, growth and respiratory metabolism of juvenileR[J]. Freshwater Fisheries, 2010, 40(2): 24-29. (in Chinese with English abstract)
[8] Thorarensen H, Gústavsson A, Mallya Y, et al. The effect of oxygen saturation on the growth and feed conversion of Atlantic halibut (L.)[J]. Aquaculture, 309(1): 96-102.
[9] Randall D J, Tsui T K. Ammonia toxicity in fish[J]. Marine Pollution Bulletin, 2002, 45(1): 17-23.
[10] Paust L O, Foss A, Imsland A K. Effects of chronic and periodic exposure to ammonia on growth, food conversion efficiency and blood physiology in juvenile Atlantic halibut (L.)[J]. Aquaculture, 2011, 315(3/4): 400-406.
[11] Foss A, Evensen T H, Vollen T, et al. Effects of chronic ammonia exposure on growth and food conversion efficiency in juvenile spotted wolffish[J]. Aquaculture, 2003, 228(1): 215-224.
[12] El-Shafai S A, El-Gohary F A, Nasr F A, et al. Chronic ammonia toxicity to duckweed-fed tilapia ()[J]. Aquaculture, 2004, 232(1): 117-127.
[13] Jensen F B. Nitrite disrupts multiple physiological functions in aquatic animals[J]. Comparative Biochemistry & Physiology Part A Molecular & Integrative Physiology, 2003, 135(1): 9-24.
[14] Siikavuopio S I, Saether B S. Effects of chronic nitrite exposure on growth in juvenile Atlantic cod,[J]. Aquaculture, 2006, 255(1): 351-356.
[15] Kroupova H, Machova J, Piackova V, et al. Effects of subchronic nitrite exposure on rainbow trout ()[J]. Ecotoxicology and Environmental Safety, 2008, 71(3): 813-820.
[16] 余瑞兰,聂湘平,魏泰莉,等. 分子氨和亚硝酸盐对鱼类的危害及其对策[J]. 中国水产科学,1999,6(3):73-77.
Yu Ruilan, Nie Xiangping, Wei Taili, et al. Toxicity of molecular ammonia & nitrite to fishes and the control measures[J]. Journal of Fishery Sciences of China, 1999, 6(3): 73-77. (in Chinese with English abstract)
[17] 张美彦,杨星,杨兴,等. 微孔曝气增氧技术应用现状[J]. 水产学杂志,2016,29(4):48-50.
Zhang Meiyan, Yang Xing, Yang Xing, et al. Current situation of application of micro-pore aeration[J]. Chinese Journal of Fisheries, 2016, 29(4): 48-50. (in Chinese with English abstract)
[18] El-Sayed A F M , Kawanna M . Optimum water temperature boosts the growth performance of Nile tilapia () fry reared in a recycling system[J]. Aquaculture Research, 2010, 39(6): 670-672.
[19] 周明,江惠龙,段玉芳,等. 非离子氨及计算方法研究[J]. 中国环境监测,1992,8(5):37-40.
[20] Abdel-Tawwab M, Hagras A E, Elbaghdady H A M, et al. Effects of dissolved oxygen and fish size on Nile tilapia,(L.): Growth performance, whole-body composition, and innate immunity[J]. Aquaculture International, 2015, 23(5): 1261-1274.
[21] 黄厚见. 摄食水平、氨氮胁迫对梭鱼幼鱼生长的影响及其毒理效应研究[D]. 上海:上海海洋大学,2012.
Huang Houjian. The Effects of Ammonia and Ration Size on Growth and Toxicological Response of Mullet,[D]. Shanghai: Shanghai Ocean University, 2012. (in Chinese with English abstract)
[22] 王甜,杜劲松,高攀,等. 氨氮对白斑狗鱼幼鱼的急性毒性研究[J]. 水产学杂志,2010,23(3):37-39.
Wang Tian, Du Jingsong, Gao Pan, et al. The toxic effect of ammonia and nitrogen on the larve of[J]. Chinese Journal of Fisheries, 2010, 23(3): 37-39. (in Chinese with English abstract)
[23] Thurston R V, Russo R C, Vinogradov G A. Ammonia toxicity to fishes. Effect of pH on the toxicity of the unionized ammonia species[J]. Environmental Science & Technology, 1981, 15(7): 837-840.
[24] 陈在新,王文一. 影响鱼类生长的水质因子机理与控制[J]. 畜牧与饲料科学,2009,30(1):15-17.
Chen Zaixin, Wang Wenyi. Mechanism and control of water quality factors influencing the growth of fish[J]. Journal of Animal Husbandry and Feed Sciences, 2009, 30(1): 15-17. (in Chinese with English abstract)
[25] García B G, Valverde J C, Aguado-Giménez F, et al. Effect of the interaction between body weight and temperature on growth and maximum daily food intake in sharpsnout sea bream ()[J]. Aquaculture International, 2011, 19(1): 131-141.
[26] 于淼. 拥挤胁迫对鱼类影响研究进展[J]. 安徽农业科学,2008,36(3):1078-1080,1082.
Yu Miao. Research progress in the effect of crowding stress on fish[J]. Journal of Anhui Agricultural Sciences, 2008, 36(3): 1078-1080, 1082. (in Chinese with English abstract)
[27] 许品诚,曹萃禾. 溶氧、水流与鱼类生长关系的探讨[J]. 淡水渔业,1989(5):27-28.
[28] 郑乐云,杨求华,黄种持,等. 循环水养殖密度和氨氮对斜带石斑鱼生长和免疫力的影响[J]. 上海海洋大学学报,2013,22(5):706-712.
Zheng Leyun, Yang Qiuhua, Huang Zhongchi, et al. Effect of breeding density and ammonia-nitrogen on growth and immunity ofin recirculating system[J]. Journal of Shanghai Ocean University, 2013, 22(5): 706-712. (in Chinese with English abstract)
[29] 罗国芝,刘刚,谭洪新. 半封闭循环水养殖系统中高体革鯻养殖密度研究[J]. 上海海洋大学学报,2012,21(2):218-224.
Luo Guozhi, Liu Gang, Tan Hongxin. The stocking density ofjuveniles in the semi-recirculating aquaculture system[J]. Journal of Shanghai Ocean University, 2012, 21(2): 218-224. (in Chinese with English abstract)
[30] 王二龙,高启平,邓玉平,等. 池塘水温溶氧变化规律及其对养殖鱼体生长的影响[J]. 科学养鱼,2017(6):21-23.
Response model construction of fish growth to water environmental factors in recirculating aquaculture mode
Cheng Xiangju1, Ju Jiaqi1, Hu Jiachun2, Xie Jun3, Yu Deguang3
(1.510641,; 2.510610,; 3.510380,)
With the improvement of aquaculture scale, the disadvantages of traditional aquaculture mode are constantly emerging, including waste and pollution of water sources. On this basis, the recirculating aquaculture system with aeration plug-flow device in pond is an exploration of efficient and energy-saving aquaculture mode, which has been developed rapidly at home and abroad. Water environment, as the place where fish live, is crucial to the growth of fish. A large number of studies have shown that dissolved oxygen, ammonia nitrogen and nitrite are the three important factors that affect the growth of fish. There are synergistic effects among dissolved oxygen, ammonia nitrogen and nitrite. Insufficient dissolved oxygen and high mass concentrations of non-ionic ammonia and nitrite can cause metabolic dysfunction of fish body and decrease of immunity, leading to the occurrence of fish diseases. Therefore, in order to explore the response of fish growth to water environment, and predict the growth rate of fish under the synergistic effect of water environment factors, the experiment was conducted on indoor recirculating aquaculture tilapia test with aerated plug-flow device with a continuous duration of 8 weeks. By changing the aeration flow, three conditions were set up in the experiment to observe the growth of tilapia during the breeding period, and the mass of the fish and water quality parameters such as dissolved oxygen, non-ionic ammonia and nitrite were regularly measured. The results showed that within a certain range, with the increase of dissolved oxygen mass concentration, the food conversion efficiency and specific growth rate of fish increased; with the increase of non-ionic ammonia mass concentration, the food conversion efficiency and specific growth rate of fish decreased. However, due to the small change in nitrite mass concentration and the safe mass concentration range, there was no significant effect on the food conversion efficiency and specific growth rate of fish in this experiment. Based on the results of the experiment, the specific growth rate of tilapia was nonlinearly fitted, and the growth prediction equation of fish was established, with2of 0.82. The validity and universality of the equation were verified by the measured data. The growth prediction equation showed that the increase of the initial mass of fish, stocking density, non-ionic ammonia and nitrite concentration could slow down the growth rate of the fish; while increasing the dissolved oxygen concentration could increase the growth rate of the fish. In the current situation of increasingly tight land, intensive farming is often adopted to improve the level of farming. With the increase of aquaculture density, the concentration of non-ionic ammonia and nitrite in the water environment increases, which have a negative impact on the growth rate of fish. It could improve the water environment and promote the healthy growth of fish by increasing dissolved oxygen. Although this prediction model is obtained under the aeration push flow cycle cultivation mode, it is also applicable to the other aquaculture water modes, making the response of fish growth to measurable water environmental factors, which provides convenience and reference for promoting the healthy development of aquaculture fish, the optimization of aquaculture system and the improvement of aquaculture benefits.
aquaculture; fish; growth; water environment; prediction model; nonlinear fitting
2018-10-30
2019-04-30
国家自然科学基金“中小型受污染水库滞水层增氧机理及效应研究”(51579106);2018年促进经济发展专项资金(现代渔业发展用途)省级项目(粤渔2018-07)
程香菊,教授,博士,主要从事湖泊、水库、河流、河口、水产养殖池塘等水力与水环境相关研究。Email:chengxiangju@scut.edu.cn:
10.11975/j.issn.1002-6819.2019.11.022
S9
A
1002-6819(2019)-11-0188-07
程香菊,具家琪,胡佳纯,谢 骏,余德光. 循环水养殖模式下鱼生长对水环境因子的响应模型构建[J]. 农业工程学报,2019,35(11):188-194. doi:10.11975/j.issn.1002-6819.2019.11.022 http://www.tcsae.org
Cheng Xiangju, Ju Jiaqi, Hu Jiachun, Xie Jun, Yu Deguang. Response model construction of fish growth to water environmental factors in recirculating aquaculture mode[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2019, 35(11): 188-194. (in Chinese with English abstract) doi:10.11975/j.issn.1002-6819.2019.11.022 http://www.tcsae.org
