甜玉米苞叶与农艺性状的遗传及相关分析
2019-07-22张丽萍谢富贵冯迪南宫捷冯发强宫庆友
张丽萍 谢富贵 冯迪南 宫捷 冯发强 宫庆友

摘要:以30份甜玉米(Zea mays L. sacharata Sturt)自交系及其配制的26個杂交组合为材料,利用加性-显性遗传模型对7个主要农艺性状和9个苞叶性状进行了遗传分析。结果表明,除最长苞叶宽外,所有性状的组合间均方都达显著或极显著差异,大多数性状的加性方差占比大于显性方差占比,所有性状的误差方差均达显著或极显著水平;对亲本的农艺性状和苞叶性状的加性效应和显性效应值进行了预测,亲本M06的多数性状表现为显著或极显著的加性正效应,组合M06×S85的多数性状表现为显著或极显著的显性正效应;相关分析发现鲜穗重分别与净穗重、穗上叶片数、穗位叶宽、苞叶干重、苞叶总面积和苞叶平均宽呈显著或极显著正相关,净穗重分别与穗位叶宽和苞叶平均宽呈显著正相关,穗位叶长和苞叶总面积、穗位叶宽与苞叶干重呈极显著正相关。
关键词:甜玉米(Zea mays L. sacharata Sturt);苞叶性状;农艺性状;遗传模型;相关分析
中图分类号:S513 文献标识码:A
文章编号:0439-8114(2019)11-0012-06
DOI:10.14088/j.cnki.issn0439-8114.2019.11.003 开放科学(资源服务)标识码(OSID):
Genetics and correlation analysis of husks and agronomic traits in sweet corn
ZHANG Li-ping1,XIE Fu-gui1,FENG Di-nan1,GONG Jie1,FENG Fa-qiang1,GONG Qing-you2
(1.College of Agriculture,South China Agricultural University,Guangzhou 510642,China;
2.Zhuhai Agricultural Science Research Center,Zhuhai 519000,Guangdong,China)
Abstract: 30 sweet corn inbred lines and 26 crosses from them were selected as the materials, 7 agronomic traits and 9 husk traits were identified and analyzed by additive-dominant genetic model. The results showed that the mean squares of all traits except for the width of the longest husk reached significant difference at the 0.05 or 0.01 level. The additive variance ratio was greater than the dominant variance ratio in most traits, and the error variance ratio reached significant at the 0.05 or 0.01 level in all traits. The additive and dominant effect values of agronomic traits and husk traits were predicted in 30 sweet corn inbred lines. The additive effect values of most agronomic traits and husk traits in M06 were positive and significant at the level of 0.05 or 0.01. The dominant effect values of most agronomic traits and husk traits in M06×S85 were positive and significant at the level of 0.05 or 0.01. The correlation analysis showed that fresh weight of ear was positively correlated with net weight of ear, number of leaves above ear, the width ear leaf of dry weight of husks, total area of husks and average width of husks, respectively. Net ear weight was positively correlated with the width of ear leaf and average width of husks, respectively. The correlations between the length of ear leaf and the total area of husks, between the width of ear leaf and dry weight of husks were positively and significant.
Key words: sweet corn(Zea mays L. sacharata Sturt); husk traits; agronomic traits; genetic model; correlation analysis
甜玉米(Zea mays L. sacharata Sturt)属于禾本科玉蜀黍族玉蜀黍属,是栽培玉米中的甜质型[1]。甜玉米种植面积逐年增长,据统计,2014年中国甜玉米种植面积达33.3万hm2,总产达499.5万t,主栽地区集中在广东、海南、广西、云南等省(自治区)[2]。随着人民生活水平的提高,对甜玉米的品质提出了更高的要求,甜玉米的品质包含外观品质、营养品质和加工品质[3,4],苞叶的颜色和形态是外观品质的重要组成部分[5]。
甜玉米的品质性状直接影响甜玉米的口感,研究最为深入,已经有大量的相关报道[6-9]。农艺性状会直接影响田间操作、产量及甜玉米果穗的外观,因此也受到较多的关注[10-12]。和凤美等[11]以超甜玉米为材料,对穗位高等12个性状进行了主成分分析,表明7个主成分可解释85%以上的表型变异。陈荣丽等[13]对甜玉米农艺性状和产量性状进行了灰色关联分析。玉米苞叶性状的研究相对较少[14-16],苞叶属于变态叶[14,17],玉米果穗苞叶能制造和贮藏子粒充实所需要的同化产物[18],还能维持子粒灌浆期间果穗的适宜温度和水分,玉米苞叶还在鲜食玉米贮运过程中具有重要作用[19]。上述的研究大多将苞叶和农艺性状分开研究,并未深入探讨两者之间的关系。
传统利用双列杂交和多系杂交进行遗传分析时,一般要求平衡数据,对缺失数据的处理难度较大。朱军[20,21]发展了基于线性模型的一系列遗传模型和统计方法,可以无偏分析不规则缺失的非平衡数据,已经在多个作物上得到了应用[22-24]。本研究采用加性-显性遗传模型,分析了30个自交系及其配制的26个杂交组合的农艺性状和苞叶性状的遗传资料,以期为甜玉米外观品质的选育及性状选择提供理论指导。
1 材料与方法
1.1 材料
选用华南农业大学甜玉米遗传育种实验室选育的30份甜玉米自交系作为亲本(表1),按多系杂交法配制26个杂交组合。2016年春播,将26个杂交组合种植于华南农业大学甜玉米遗传育种实验室玉米试验地,随机区组排列,重复3次,2行区,每行10株,株距40 cm,行距60 cm,农事操作与大田生产相同。
1.2 方法
每小区于授粉后第22天,取生长相对一致的连续5株测量农艺性状和苞叶性状,取平均值进行数据分析。测量的农艺性状为鲜穗重、净穗重、穗长、穗行数、穗上叶片数、穗位叶长和穗位叶宽;苞叶性状为苞叶层数、苞叶干重、苞叶总面积、苞叶平均长、苞叶平均宽、最外苞叶长、最外苞叶宽、最长苞叶长和最长苞叶宽,测量方法参考盖钧镒[25]、吕桂华等[15]。苞叶干重以120 ℃烘干至恒重为计;苞叶面积=0.75×苞叶长×苞叶宽。
采用SAS软件进行方差分析,参照加性-显性遗传模型进行农艺性状和苞叶性状遗传参数的分析。采用QGAStation(http://ibi.zju.edu.cn/software/qga/)分析軟件估算各项方差分量占表型方差的比率,估算成对性状间的相关系数、表型相关系数和各项遗传相关系数,采用调整无偏预测法(AUP)预测各项遗传效应值,用以基因型为抽样单位的Jackknife抽样方法估算各项统计量的标准误,之后检测各遗传参数的显著性。
2 结果与分析
2.1 农艺性状和苞叶性状方差分析
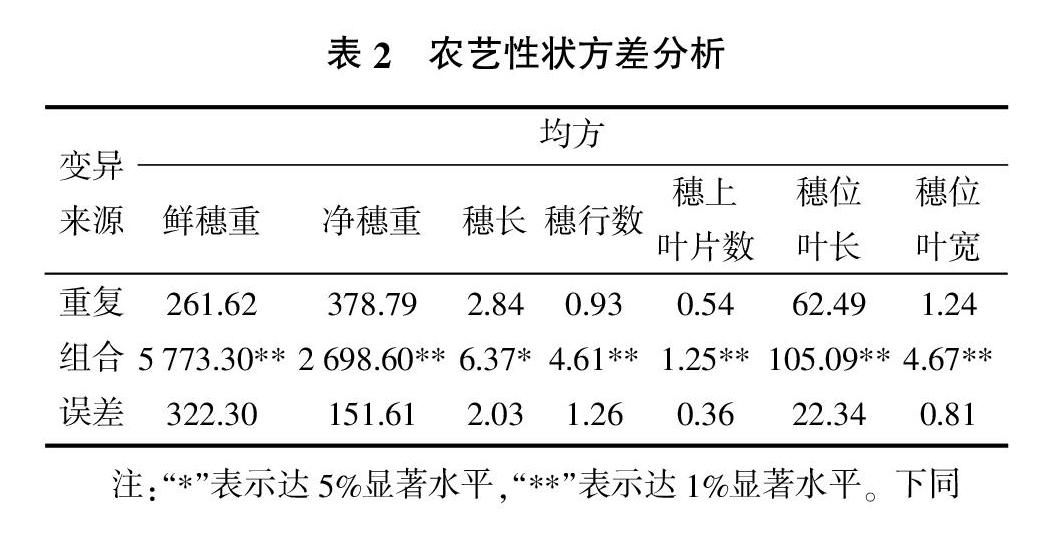
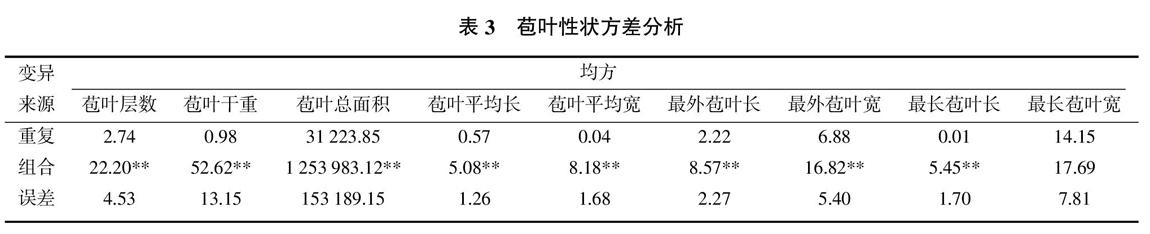
对30个亲本自交系及其配制的杂交组合的农艺性状和苞叶性状进行方差分析,结果表明,所检测的农艺性状基因型间的均方均达显著或极显著水平;所检测的9个苞叶性状中,除最长苞叶宽外,其余性状均达极显著水平(表2、表3)。说明这些性状在不同基因型间存在较大的变异范围及显著的差异,可进一步进行遗传分析。
2.2 农艺性状和苞叶性状的遗传方差组成
甜玉米7个农艺性状的各项遗传方差分量占表型方差的比率见表4。穗行数、穗上叶片数和穗位叶宽的加性方差占表型方差的比率均为显著或极显著水平,但所占比重有所不同。仅穗位叶长的显性方差占表型方差的比率为显著水平,大部分性状的加性方差比率高于显性方差比率,表明农艺性状以加性基因控制为主。研究的农艺性状的随机误差方差比率均超过了30%,且达到了显著或极显著水平,说明农艺性状受环境和随机误差的影响较大。
9个苞叶性状的各项遗传方差占表型方差的比率见表5。苞叶层数、苞叶干重、苞叶总面积、苞叶平均宽和最长苞叶宽的加性方差占表型方差的比率达显著或极显著水平,仅苞叶总面积的显性方差占表型方差的比率达显著水平,说明苞叶性状主要由加性效应所决定。所有苞叶性状的随机误差方差比率均超过了30%,均达显著或极显著水平,表明苞叶性状受环境和随机误差的影响也较大。
从性状分析,穗行数、穗上叶片数、穗位叶宽和苞叶层数这4个性状的加性方差占表型方差的比率较高,可通过改良自交系来改良杂交种的表型。穗位叶长和苞叶总面积的显性方差占表型方差的比率达显著水平,可通过选配杂交组合,利用特定环境下的杂种优势来改良表型。随机误差方差占表型方差的比率较高的性状为最外苞叶长、最外苞叶宽、鲜穗重、净穗重和穗长,表明这些性状的环境互作十分重要,性状的改良受环境的影响较大。
2.3 亲本农艺性状和苞叶性状的加性效应
根据遗传方差分析的结果,农艺性状中除穗长、穗位叶长外,其余性状均加性方差大于显性方差;苞叶性状中除苞叶总面积、最外苞叶长和最外苞叶宽外,其余性状也是加性方差大于显性方差,可对亲本的加性效应进一步分析。30个亲本的农艺性状和苞叶性状的加性效应预测值见表6和表7。其中M06、M08、M14和S63的鲜穗重、净穗重和穗位叶宽均表现为显著或极显著水平的正效应,可用这3份材料为亲本改良后代这些性状的表现;M06的苞叶性状加性效应值也大多表现为显著或极显著的正效应,可用于亲本改良后代的这些性状。
各农艺性状正加性效应值最高的前2位亲本:鲜穗重为M08和M06,净穗重为M14和M08,穗长为S63和M10,穗行数为S85和F106,穗上叶片数为M06和S83,穗位叶长为M10和D53,穗位叶宽为M06和S63。各苞叶性状正加性效应值最高的前2位亲本:苞叶层数为M06和S85,苞叶干重为M10和S68,苞叶总面积为M06和S85,苞叶平均长为D53和S75,苞叶平均宽为M10和M08,最长苞叶长为D53和S75,最长苞叶宽为M06和M08。
2.4 杂交组合农艺性状和产量性状的显性效应
26个杂交组合的农艺性状和苞叶性状的显性效应预测值见表8。有3个农艺性状,即穗行数、穗上叶片数和穗位叶宽未检测到显性效应,说明这些性状受显性效应的影响较小。组合M06×S85除最外苞叶宽外,其他性状均表现为正效应值,且有5个性状显性效应达显著或极显著水平,说明这个组合的杂种优势最大;组合M08×S83的鲜穗重和净穗重表现为显著正效应,也存在较大的杂种优势。
2.5 农艺性状和苞叶性状间的相关分析
对甜玉米的7个农艺性状和9个苞叶性状进行相关分析,结果(表9)表明,鲜穗重分别与净穗重、穗上叶片数、穗位叶宽、苞叶干重、苞叶总面积和苞叶平均宽呈显著或极显著正相关,净穗重分别与穗位叶宽和苞叶平均宽呈显著正相关,未检测到穗长和穗行数与其他性状间的显著相关性。检测到穗上叶片数与苞叶干重和苞叶总面积间呈极显著正相关,相关系数分别为0.66和0.67;穗位叶长和苞叶总面积间呈极显著正相关,相关系数为0.71;穗位叶宽与苞叶干重呈极显著正相关,相关系数为0.75。9个苞叶性状间检测到8对性状间呈显著或极显著相关,分别为苞叶层数与苞叶总面积、苞叶干重与苞叶总面积和苞叶平均宽、苞叶平均长与最外苞叶长和最长苞叶长、苞叶平均宽与最外苞叶宽和最长苞叶宽、最外苞叶长与最长苞叶长,且相关系数均为正值。
3 讨论
甜玉米的农艺性状和苞叶性状,如鲜穗重、净穗重、穗长等都属于数量性状,受微效多基因控制,并且易受环境条件的影响。以往对这类性状进行遗传分析,大多采用双列杂交的分析方法估算一般和特殊配合力[26-29],通过计算一般配合力方差与特殊配合力方差的比值来分析性状主要受何种效应控制。传统的分析方法未考虑到基因与环境的互作,且无法分析多系杂交且有不规则缺失的遗传资料,采用混合线性模型的加性-显性模型[30,31]可以估算遗传方差的同时预测遗传效应值。
本研究发现甜玉米的多数农艺性状和苞叶性状的加性方差大于显性方差,且多数性状受环境因子的影响较大,与前人的结果一致。蔡一林等[32]对玉米株型遗传分析发现叶向值、雄穗分枝数符合加性-显性遗传模型,叶面积符合加性-上位性遗传模型。黄成威等[33]对甜玉米保鲜性状研究发现含糖量下降速率受加性-显性-上位性主基因和加性-显性多基因遗传控制。霍仕平等[34]以双列杂交的方法研究发现果穗苞叶数、苞叶长、苞叶宽等性状符合加性-显性遗传模型。吕桂华等[15]研究发现苞叶层数、最长苞叶长、鲜苞叶重和干苞叶重受加性基因效应控制,苞叶厚度、最外苞叶长(宽)、最长苞叶宽、最内苞叶长(宽)以加性效应为主,存在显性和上位性效应。玉米是异花授粉作物,重视杂种优势的利用,因此在杂交组合选配时要注意结合特定的生态条件进行。
苞叶是果穗营养的贮存器官,也是果穗发育的保护器官,对普通玉米而言,苞叶层数较多或较厚不利于子粒成熟后的脱水[18],而苞叶性状是甜玉米外观品质的一个重要方面[1],完整且紧密的苞叶有利于阻止害虫对甜玉米果穗的危害,增加外观得分。本研究检测到苞叶层数、苞叶平均宽和最长苞叶宽的加性方差比率较高,可在早代进行选择,而苞叶总面积的显性方差比率较高,选育时应充分考虑杂种优势的利用。本研究中也检测到鲜穗重分别与净穗重、穗上叶片数、穗位叶宽、苞叶干重、苞叶总面积和苞叶平均宽呈显著或极显著正相关,净穗重分别与穗位叶宽和苞叶平均宽呈显著正相关,可见加强对甜玉米苞叶干重、总面积和苞叶宽的选择有利于提高子粒产量。
参考文献:
[1] 刘纪麟.玉米育种学[M].北京:农业出版社,1991.
[2] 刘蔚楠,万 忠,甘阳英,等.2015年广东甜玉米产业发展形势与对策建议[J].广东农业科学,2016(3):12-16.
[3] 王 娜,史振声,王志斌,等.甜玉米品质研究进展[J].玉米科学,2007(6):47-50.
[4] 王振华.甜玉米品质性状与部分农艺性状的相关分析[J].玉米科学,1998(2):22-25.
[5] TRACY W F,HALLAUER A R. Specialty corns[J].World vegetables,1994,81(11):777-807.
[6] ZHANG S L,HUANG Y Q,HE Z H,et al. Variation law of pericarp tenderness of super sweet corn kernel[J].Agricultural science & technology,2016,17(7):1671-1674.
[7] 馮发强,王国华,王青峰,等.甜玉米类胡萝卜素合成关键基因PSY1、LCYE和CrtRB1的功能分析[J].华南农业大学学报,2015,36(5):36-42.
[8] 于永涛,李高科,祁喜涛,等.甜玉米果皮厚度QTL的定位及上位性互作[J].作物学报,2015,41(3):359-366.
[9] 张姿丽,蒋 锋,刘鹏飞,等.鲜食甜玉米籽粒蛋白质含量的QTL定位[J].西北农林科技大学学报(自然科学版),2013,41(3):99-104.
[10] 张姿丽,蒋 锋,刘鹏飞,等.甜玉米穗位叶面积QTL定位[J].湖北农业科学,2014,53(7):1502-1505.
[11] 和凤美,朱永平,朱 芮,等.超甜玉米自交系主要农艺性状及鲜穗产量的主成分分析[J].中国农学通报,2014,30(18):79-83.
[12] 郭惠明,王志纯,赖伦英.福建甜玉米区试品种产量及农艺性状分析[J].亚热带农业研究,2013,9(2):78-81.
[13] 陈荣丽,宋文兰,周 胜,等.甜玉米鲜穗产量与农艺性状灰色关联度分析[J].种子,2018,37(9):92-95.
[14] 马智艳,董永彬,乔大河,等.不同种质玉米杂交种苞叶性状特征分析[J].河南农业科学,2015,44(2):15-18.
[15] 吕桂华,陈坚剑,徐秀红,等.鲜食糯玉米苞叶相关性状的遗传相关分析[J].浙江农业学报,2015,27(7):1122-1126.
[16] 孟剑霞.玉米雌穗与苞叶发育研究进展[J].安徽农学通报,2007, 13(14):78-79.
[17] 宋凤斌,徐洪文.玉米苞叶光合生理特性研究进展[J].玉米科学,2008,16(4):31-34.
[18] CUI Z,LUO J,QI C,et al. Genome-wide association study (GWAS) reveals the genetic architecture of four husk traits in maize[J].BMC genomics,2016,17(1):946.
[19] CAO A,SANTIAGO R,RAMOS A J,et al. Critical environmental and genotypic factors for Fusarium verticillioides infection,fungal growth and fumonisin contamination in maize grown in northwestern Spain[J].International journal of food microbiology,2014,177(3):63-71.
[20] 朱 军.遗传模型分析方法[M].北京:中国农业出版社,1997.
[21] 朱 军.广义遗传模型与数量遗传分析新方法[J].浙江农业大学学报,1994,20(6):551-559.
[22] 阮仁武,傅大雄,戴秀梅.小麦主要产量性状的杂种优势和遗传分析[J].西南大学学报(自然科学版),2002,24(2):141-145.
[23] 肖炳光,朱 军,卢秀萍,等.烤烟主要农艺性状的遗传与相关分析[J].遗传,2006,28(3):317-323.
[24] 陈建国,朱 军.籼粳杂交稻米外观品质性状的遗传及基因型×环境互作效应研究[J].中国农业科学,1998,31(4):1-7.
[25] 盖钧镒.作物育种学各论[M].北京:中国农业出版社,2006.
[26] 刘艳改.陆地棉农艺与纤维品质性状的双列杂交分析[D].北京:中国农业科学院,2014.
[27] 刘春晓,赵海军,董树亭,等.玉米不同基因型双列杂交后代抽丝后氮素代谢特性[J].中国农业科学,2014,47(1):33-42.
[28] 贺建波,管荣展,盖钧镒.双列杂交设计的主-微位点组遗传分析方法研究[J].作物学报,2010,36(8):1248-1257.
[29] 沈法富,于元杰,毕建杰,等.棉花耐盐性的双列杂交分析[J].作物学报,2001,24(1):50-54.
[30] 朱 军.作物杂种后代基因型值和杂种优势的预测方法[J].生物数学学报,1993,8(1):32-44.
[31] 朱 軍.估算遗传方差和协方差的混合模型方法[J].生物数学学报,1992,7(1):1-11.
[32] 蔡一林,王久光,孙海燕,等.玉米几个株型性状的遗传模型及其与穗粒性状的典型相关分析[J].作物学报,2002,25(6):829-834.
[33] 黄成威,陈沛君,秦彤非,等.甜玉米保鲜性状的遗传模型研究[J].湖北农业科学,2016,55(6):1375-1378.
[34] 霍仕平,晏庆九,许明陆,等.玉米果穗苞叶性状的遗传分析[J].杂粮作物,2000,20(2):8-12.
