大丁草属2种植物花粉萌发与贮藏特性研究
2019-07-22刘春雪李叶芳关文灵
刘春雪 李叶芳 关文灵


摘要:以野生大丁草、钩苞大丁草花粉为材料,采用离体培养法,研究大丁草属2种植物花粉萌发的最适条件以及适合花粉保存的贮藏方法。结果表明,大丁草和钩苞大丁草花粉萌发最适宜的培养基相同,均为部分ME3+600 g/L PEG200,最高萌发率分别为47.11%和44.40%。2种植物花粉萌发最适合的温度差别较大,最适合大丁草花粉萌发的温度为30 ℃,萌发率为51.09%;而最适合钩苞大丁草花粉萌发的温度为25 ℃,萌发率为51.90%。2种植物花粉自然条件下的离体寿命均较短,仅可维持48 h。4 ℃低温干燥贮藏有利于延长2种植物花粉的寿命,贮藏15 d后萌发率均高于10%,而室温贮藏、-20 ℃冷冻贮藏15 d后萌发率均低于10%。
关键词:大丁草属;花粉萌发力;适宜条件;离体寿命;贮藏方式
中图分类号: Q945.34 文献标志码: A 文章编号:1002-1302(2019)11-0173-04
收稿日期:2018-02-03
基金项目:云南省现代农业花卉苗木产业技术体系建设项目(编号:2017KJTX0010)。
作者简介:刘春雪(1992—),女,河北秦皇岛人,硕士研究生,主要从事园林植物资源利用与创新研究。E-mail:306143396@qq.com。
通信作者:关文灵,博士,教授,主要从事园林植物种质资源及植物造景研究。E-mail:158066692@qq.com。 大丁草属(Gerbera)植物是菊科(Compositae)帚菊木族中的一属,全世界有近80种,主要分布于非洲,次为亚洲东部及东南部。我国有20种,除个别种遍及于南北各地外,绝大部分集中于西南地区,云南有16种,占全国种类的80%[1]。
该属最常见的植物非洲菊(Gerbera jamesonii)观赏价值和经济价值较高,是一种应用广泛的切花材料[2]。本属其他植物在国内多作民间用药[3],如大丁草(Gerbera anandria)可用于清热利湿、解毒消肿、止咳、止血[4];钩苞大丁草(Gerbera delavayi)可清热利湿、消积杀虫,也可作为我国西南少数民族人民编织火草服饰的原料[5-6]。
大丁草和钩苞大丁草为大丁草属的2个野生种。经过野外调查以及田间观察,2个种对白粉病抗性较强。将大丁草、钩苞大丁草与非洲菊进行种间远缘杂交,有望培育出抗病性较强的非洲菊新品种。因此需要充分了解大丁草、钩苞大丁草花粉生活力和贮藏特性。目前,对大丁草、钩苞大丁草的研究主要集中于化学成分及药理活性[3-4,7]、环境胁迫对种子萌发的影响[8-10]、组织培养及快繁技术[11-12]、遗传多样性[5-6,13]、基因测序分析[14-15]等,花粉萌发方面的研究未见报道。本试验以野生大丁草、钩苞大丁草为材料,采用液体培养基萌发法来筛选适合大丁草花粉萌发的培养基和培养温度,并探讨不同贮藏条件对大丁草、钩苞大丁草花粉生活力的影响,旨在为其远缘杂交提供依据。
1 材料与方法
1.1 材料
于2015年11月从云南省禄丰县采集生长健壮的野生大丁草、钩苞大丁草种苗,并栽于云南农业大学园林苗圃大棚内。栽植株行距为16 cm×16 cm,栽后进行常规田间管理。大丁草于2016年9月开始陆续开花,钩苞大丁草于2016年12月开始陆续开花。于晴天08:00—09:00采集盛花期花粉。
1.2 试验时间及地点
试验于2016年9月至2017年1月在云南农业大学园林园艺学院实验室进行。
1.3 试验方法
采用离体萌发法[16],花粉萌发培养基参考赵宏波等的方法[17],以部分ME3培养基(MgSO4·7H2O 370 mg/L、KNO3 950 mg/L、CaCl2·2H2O 880 mg/L、H3BO3 50 mg/L)为基础,将液体培养基滴于凹面载玻片,把花粉散播于培养基,放于装有湿滤纸的培养皿中,根据试验需要培养。花粉离体萌发率统计以花粉管长于花粉直径为标准,每次每个处理分别取3个固定大小的视野统计,单次统计的花粉数不少于50个,重复3次[2]。
1.4 试验设计
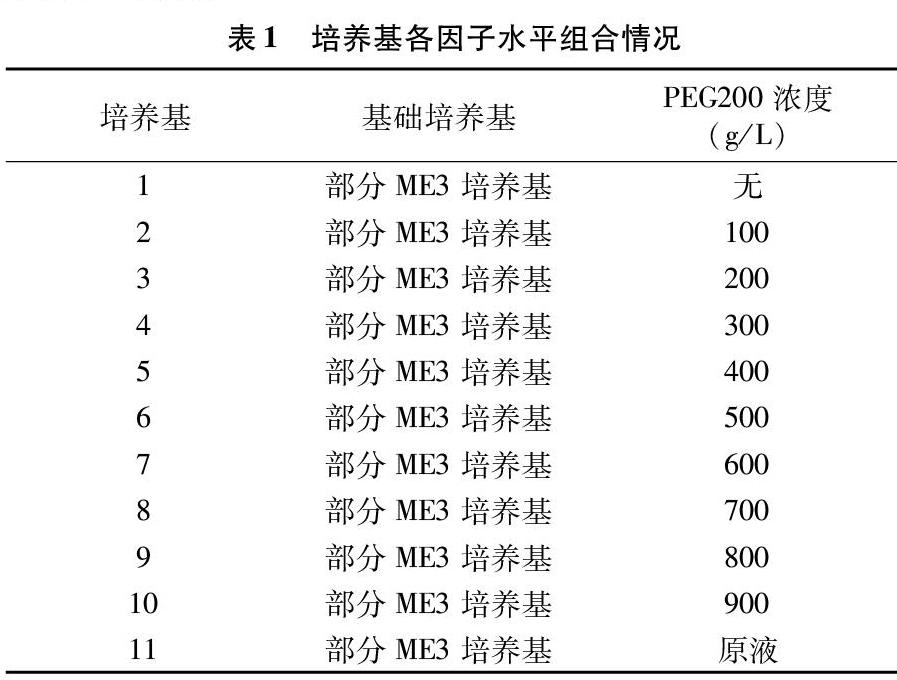
1.4.1 不同浓度的PEG200对花粉萌发的影响 采集新鲜的大丁草、钩苞大丁草花粉,分别撒播在添加了不同浓度的PEG200的部分ME3培养基上(表1),置于25 ℃的智能光照培养箱中培养。24 h后在生物显微镜下观察每种培养基上花粉萌发的情况。根据结果筛选出最适于花粉萌发的PEG200浓度。
1.4.2 花粉萌发最适温度筛选 采集新鲜大丁草、钩苞大丁草花粉,使用“1.4.1”节所得的最适培养基,在不同温度的智能光照培养箱中培养24 h后观察不同温度对花粉萌发的影响,设置处理温度为15、20、25、30、35、40 ℃。统计并计算萌发率,根据结果筛选出最适合花粉萌发的温度。
1.4.3 贮藏方法对花粉寿命的影响 采集新鲜大丁草、钩苞大丁草花粉,不经过任何处理放于室温下,分别于0、1、2、4、6、12、24、30、48 h后测定花粉活力,培养24 h后观察花粉萌发情况。使用“1.4.1”节所得的最适培养基,“1.4.2”节所得的最适温度。记录并统计萌发率(萌发率=萌发的花粉数/总的花粉数×100%)。
测定出室温条件下花粉的寿命。同时测定贮藏方法对花粉寿命的影响。采集新鲜大丁草、钩苞大丁草花粉,首先放置于装有硅胶干燥剂的干燥器内干燥24 h,然后分三部分贮藏:(1)室温贮藏。将干燥后的花粉用封口膜封口,放入密闭、装有硅胶干燥剂的自封袋中,将自封袋封口,置于室温环境贮藏。(2)4 ℃冷藏贮藏。将干燥后的花粉用封口膜封口,放入密闭、装有硅胶干燥劑的自封袋中,将自封袋封口,置于4 ℃冰箱内冷藏贮藏。(3)-20 ℃冷冻贮藏。将干燥后的花粉用封口膜密封封口,放入密闭、装有干硅胶燥剂的自封袋中,将自封袋封口,置于-20 ℃冰箱内冷冻贮藏。分别在5、10、15 d 后测定不同贮藏方式下的花粉萌发力。使用“1.4.1”节所得的最适培养基,“1.4.2”节所得最适温度,记录并统计萌发率,并根据结果得出花粉最适贮藏温度。
在父本、母本花期不遇时,需对花粉进行妥善贮藏。在室温条件下,大丁草属2种植物的花粉活力均下降最快,15 d后2种植物的花粉萌发率均低于10%。对地被菊[24]、小菊花[31]等菊科植物花粉的贮藏研究也表明,室温条件不利于花粉活力的保持和寿命的延长。原因可能是室温环境下酶活性强,花粉的呼吸作用等生理代谢活动活跃,使活力下降迅速。有研究表明,低温可降低花粉呼吸作用及其他生理功能,有利于花粉较长时期保存活力[32]。尾叶紫薇花粉在干燥6~8 h后,在4 ℃条件下贮藏25 d后仍然保持较高的萌发率和较长的花粉管[33];菠萝蜜花粉在干燥3 h后,在4 ℃的环境条件下生命力可延长20 d[34]。本研究显示,大丁草属2种植物的花粉在4 ℃低温干燥贮藏条件下的活力变化趋势与其他植物一致,可有效延长花粉寿命,在贮藏15 d后2种植物的花粉活力均高于10%。-20 ℃低温冷冻条件与室温条件相比,2种植物的花粉活力也维持在较高水平,但与4 ℃储藏相比没有显著优势。因此大丁草屬2种植物花粉最好的贮藏方法是 4 ℃ 低温干燥冷藏,但贮藏时间不宜超过15 d。
影响花粉寿命的因子还有许多,如花粉含水量、干燥时间、贮藏环境的相对湿度以及超低温等[35-36],它们对大丁草属2种植物花粉生活力的影响以及更好的贮藏方法还有待于进一步研究。
参考文献:
[1]中国科学院中国植物志编辑委员会.中国植物志[M]. 北京:科学出版社,1996:73.
[2]孙 强,芦建国,沈永宝,等. 非洲菊花粉和柱头生物学习性初步研究[J]. 上海交通大学学报(农业科学版),2008,26(1):78-80,90.
[3]肖 瑛,丁 怡. 大丁草属植物的化学成分和药理活性研究[J]. 天然产物研究与开发,2002,14(6):51-57.
[4]He F,Wang M,Gao M H,et al. Chemical composition and biological activities of Gerbera anandria[J]. Molecules,2014,19(4):4046-4057.
[5]徐晓丹. 钩苞大丁草的纤维特性及遗传多样性研究[D]. 武汉:华中农业大学,2016.
[6]欧成川. 钩苞大丁草的叶片形态多样性研究及优异纤维种质发掘[D]. 昆明:昆明理工大学,2016.
[7]谷黎红,李 铣,陈英杰,等. 大丁草中抗菌活性成分的研究:人肠道微生物对大丁甙及其类似物的代谢产物[J]. 药学学报,1988(7):511-515.
[8]姜云天,李玉梅,顾地周,等. Na2CO3胁迫对野生大丁草种子萌发的影响[J]. 种子,2014,33(6):86-88.
[9]姜云天,沈红梅,顾地周,等. NaCl胁迫对长白山区野生大丁草种子萌发的影响[J]. 现代农村科技,2014(17):59-59,60.
[10]姜云天,顾地周,周 繇,等. 盐胁迫对野生大丁草种子萌发的影响[J]. 中国园艺文摘,2014(8):25-27.
[11]凌 杰,宁淑香. 大丁草愈伤组织及试管苗培养的研究[J]. 安徽农业科学,2012,40(5):2671-2673.
[12]黄玉玲,朱跃珍,李永平,等. 火草组培快繁技术研究[J]. 林业调查规划,2016,41(4):72-74,79.
[13]欧成川,郑 伟,王谈笑,等. 钩苞大丁草的叶片形态多样性研究[J]. 浙江农业科学,2016,57(2):194-197.
[14]陈 菁,郑 伟,王谈笑,等. 钩苞大丁草高通量转录组测序及差异表达分析[J]. 西北植物学报,2017,37(3):470-477.
[15]王谈笑,郑 伟,陈 菁,等. 基于ITS序列分析钩苞大丁草九个居群的亲缘关系[J]. 广西植物,2017,37(3):329-334.
[16]胡适宜. 植物学实验方法(一):花粉生活力的测定[J]. 植物学通报,1993,10(2):60-62.
[17]赵宏波,陈发棣,房伟民. 栽培小菊和几种菊属植物花粉离体萌发研究[J]. 南京农业大学学报,2005,28(2):22-27.
[18]Zhang H Q,Croes A F. A new medium for pollen germination in vitro[J]. Plant Biology,1982,31(1/2):113-119.
[19]Subbaiah C C. A polyethylene glycol based medium for in vitro germination of cashew pollen[J]. Canadian Journal of Botany,1984,62(12):2473-2475.
[20]Roberts I N,Stead A D,Ockendon D J,et al. Pollen stigma interactions in Brassica oleracea[J]. Theoretical and Applied Genetics,1980,58(6):241-246.
[21]宋红霞,吴旭艳,武 喆,等. PEG对胡萝卜花粉离体萌发的影响[J]. 长江蔬菜,2011(6):22-24.
[22]何 琪,何 芳,刘 鹏,等. PEG-4000、蔗糖及pH值对七子花花粉萌发的影响[J]. 湖北农业科学,2006,45(2):214-216.
[23]Jamesl B,Beyoungh K. The essential role of calcium ion in pollen germination and pollen tube growth[J]. American Journal of Botany,1963,50(9):859-865.
[24]吕晋慧,赵 耀,任 意,等. 地被菊花粉生活力测定及影响因素研究[J]. 华北农学报,2011,26(4):189-193.
[25]叶利民. PEG和钙对栀子花粉萌发的影响[J]. 种子,2012,31(7):35-36,39.
[26]赵剑颖,张华丽,张西西,等. 万寿菊花粉活力及柱头可授性研究[J]. 中国农学通报,2012,28(19):159-163.
[27]刘林德,张洪军,祝 宁. 刺五加花粉活力和柱头可授性的研究[J]. 植物研究,2001,21(3):375-380.
[28]姬慧娟,尹林克,严 成,等. 多枝柽柳的开花动态及花粉活力和柱头可授性研究[J]. 西北农林科技大学学报(自然科学版),2009,37(5):114-118.
[29]胡适宜. 被子植物胚胎学[M]. 北京:人民教育出版社,1982:51-58.
[30]孙 强. 非洲菊杂交育种技术及优良品系选育初步研究[D]. 南京:南京林业大学,2007.
[31]王 涛,祝朋芳,董玉芝. 小菊花粉生活力及贮藏力的研究[J]. 辽宁林业科技,2010(1):8-11.
[32]尹佳蕾,赵惠恩. 花粉生活力影响因素及花粉贮藏概述[J]. 中国农学通报,2005,21(4):110-113,193.
[33]王晓玉,徐 婉,胡 杏,等. 尾叶紫薇开花及花粉萌发研究[J]. 浙江农林大学学报,2012,29(6):966-970.
[34]余 庆,李映志,汪永保,等. 菠萝蜜花粉萌发及贮藏特性研究[J]. 广东农业科学,2014,41(8):60-64.
[35]杨 红,顾 妍,张朝阳,等. 不同辐射及温度处理对西瓜花粉生活力的影响[J]. 江苏农业科学,2017,45(2):111-113.
[36]张 婷,赵 林,张 梅,等. 不同苹果品种花粉的生物学特性[J].
