壮体沙鳅早期发育及红水河来宾江段资源补充量评估
2019-07-20高明慧吴志强黄亮亮谭细畅赵立朝
高明慧 吴志强 黄亮亮 谭细畅 郑 雄 赵立朝
(1. 桂林理工大学环境科学与工程学院, 桂林理工大学广西岩溶地区水污染控制与用水安全保障协同创新中心, 桂林 541004;2. 广西大学动物科学技术学院, 南宁 530004; 3. 珠江流域水资源保护局, 广州 510380)
壮体沙鳅[Botia robusta(Wu, 1939)], 属鲤形目,鳅科, 沙鳅亚科, 沙鳅属; 其为小型鱼类, 喜流水, 广泛分布于长江、珠江水系上、中游, 为红水河干流的优势种类[1]。鱼类早期发育是形态、生态、生理行为等方面动态变化的过程, 同时, 也是鱼类生活史中死亡率很高的时期, 这一阶段的存活率直接影响到鱼类补充量的大小[2]。对壮体沙鳅早期发育及其补充群体状况的调查研究, 不仅可丰富其生物学资料, 对其物种鉴定、资源量分析及其资源保护等也具有重要的意义, 同时可为其他鱼类的丰度及协调作用的影响研究提供基础数据。目前有关淡水鱼类早期发育的研究主要集中于常见、经济种类,如对长江流域102种鱼类的早期形态描述[3]、珠江流域西江49种常见鱼类的早期发育描述[4]及谭细畅等[5,6]对鲤(Cyprinus carpio)、赤眼鳟(Squaliobarbus curriculus)早期发育的系统研究等。随着河流生态环境的改变, 西江鱼类群落结构也随之改变,经济性鱼类资源量明显下降下降[1], 如广东鲂(Me-galobrama terminalis)[7], 小型鱼类逐渐成为优势种的趋势日渐明显。然而现有的小型鱼类早期形态发育资料相对较少, 且多没有系统的分期描述。于壮体沙鳅而言, 仅郑惠芳[8]对其生态学特征进行了研究, 曹文宣等[3]发现其产卵时间主要集中在5—6月份。
早期发育阶段鱼类的分类鉴定是鱼类早期资源调查中至关重要的一项工作。基于形态特征的传统鉴定方法极为细致繁琐, 往往长期从事分类研究的专家才能进行准确鉴定。为此人们开始寻求更为简单可靠的鉴定方法, 其中主要为计算机辅助方法和生化-遗传方法。这2种方法的优点是自动化程度高、辨别精度高、效率快, 以Froese[9]为代表, 但在采样现场对已有资料的种类辨别时或对某些同属种类而言, 形态鉴别方法较计算机辅助方法和生化-遗传方法更为简单有效[10], 且计算机辅助方法是在传统形态数据的基础上, 依据不同的数据分析模式进行判别得到结果。
由此可知, 鱼类的早期形态观察描述仍十分必要。本文对红水河壮体沙鳅的鱼卵进行了培养和观察, 对主要的发育期进行了系统的拍照和描述,此外对壮体沙鳅鱼卵的单位捕捞努力量(CUPE)及早期发育特征的种间比较和资源现状提供基础资料。
1 材料与方法
1.1 调查方法
2017年4月16日至8月30日的07:00—08:00, 于红水河来宾段的三江口(E 109°30′35″, N 23°48′28″)处用弶网(网口1.0 m×1.5、网目0.5 mm、网长6 m)、集苗箱(0.8 m × 0.4 m × 0.4 m)进行近岸定点采集(图 1)。使用LJD-10A打印式流速仪测量网口流速以计算滤水量, 水银温度计测量气温和水温, 萨氏盘测量水体透明度, 并通过查询全国水雨情信息网http://xxfb.hydroinfo.gov.cn获取水位和流量数据。
1.2 鉴别与培育
将采集到的鱼卵带回室内, 在体式解剖镜下依据卵膜颜色、卵膜性质、卵膜直径和肌节等特征对其进行种类鉴定[3,4]。对不能确定种类的鱼卵进行分类、编号, 放入50 mL的培养杯中单独培养。水温维持在22—25℃, 每日换水1次, 保证溶氧充足。胚胎孵出2d后开始投喂煮熟的蛋黄, 且0.5h后将多余的蛋黄吸出以维持水质良好, 待达到稚鱼期时作进一步鉴别[10—12]。
1.3 观察方法
对单独培养的鱼卵在体视显微镜下用尼康(Nikon)5400进行观察和照片拍摄, 卵期每30min观察拍照1次。卵黄囊期仔鱼, 每8h观察拍照1次, 之后根据发育情况逐步延长观察拍照的时间间隔。记录及拍照结束之后将仔鱼继续单独培养, 对难以活体拍照的时期, 用5%的福尔马林固定后进行拍照, 同时从单独培养的鱼卵中选取相同种类作为下一时期的描述对象。以此方法对每日采集的壮体沙鳅鱼卵进行多样本的形态学描述和统计。
1.4 数据分析
鱼苗径流量参考易伯鲁等[11]的调查方法进行,其计算公式为:

其中:M为单位时间鱼苗径流量(尾),Q为采集点断面的平均江水流量(m3/s),q为流经网内的江水流量(m3/s),m为采集到的鱼苗数量(尾),c为断面鱼苗系数取0.39。全天的鱼苗径流量用单位时间鱼苗径流量换算得出。
产卵场位置(L)的计算公式为:

T为采集到的卵或早期仔鱼的发育时间, 根据胚胎发育时序来确定;V为采样期间采样点上游江段平均流速。
壮体沙鳅鱼卵单位捕捞努力量(CUPE)与流量、水温、透明度等水文环境因子变化的相关性采用Pearson Correlation法分析, 通过SPSS20.0完成。
2 结果
2.1 壮体沙鳅早期形态
壮体沙鳅鱼卵的特征 壮体沙鳅的卵基本为规则球形, 呈淡黄色, 卵周隙较大, 卵膜最大直径可达6.8 mm。所采集鱼卵多为尾牙期, 眼基、肌节、听囊、尾鳍、心脏、胸鳍等分化形成, 肌节出现17对, 尾部开始突出。

图 1 红水河采样点图Fig. 1 Location of sampling site at Hongshuihe River
胚胎阶段尾牙期:卵膜径2.6 mm(图版Ⅰ-1), 胚层已完全下包, 具胚体雏形, 脑明显分化, 肌节17对, 尾部开始突出, 胚体整体呈淡黄色。
肌肉效应期:尾牙期后3.5h, 卵膜径为3.6 mm(图版Ⅰ-2), 胚体和卵黄囊延长, 听囊明显, 脑区发达, 隆起明显, 尾斜垂, 肌节23对, 胚体躯干开始间歇性颤动, 但频率较低。
心脏原基期:尾牙期后5.5h, 卵膜径为4.3 mm(图版Ⅰ-3), 后脑与卵黄囊连接处出现心脏原基, 胚体中部向内凹, 有节律地收缩, 卵黄囊后端随着尾部的延伸拉长, 在听囊处可见2个小点, 为耳石雏形,肌节26对。
心脏搏动期:尾牙期后11h, 卵膜径为5.2 mm(图版Ⅰ-4), 心脏开始搏动频率较低为65次/min, 卵膜软化且出现裂缝, 胚体开始间歇性的抖动, 卵黄囊后端随着尾部的延伸变直, 肌节31对。
胸鳍原基期:尾牙期后15h, 卵膜径为5.5 mm(图版Ⅰ-5), 听囊后方出现芽状胸鳍原基, 卵膜软化,肌节对32对, 胚体剧烈冲撞卵膜。
仔鱼阶段孵出期:全长5.8 mm, 肌节32对(图版Ⅰ-6); 头部尚未与卵黄囊分离; 卵黄囊呈锥形; 尾鳍褶呈现圆形; 肛门原基已形成; 背腹缘鳍褶发达; 身体透明无色素分布; 心脏位于卵黄囊前方,由心房、心室和静脉窦组成, 内部血液流动明显,心跳约 180 次/min; 仔鱼活动能力很强, 不停抖动身体。
眼黑色素期:孵出后18h, 全长为6.5 mm, 肌节32对(图版Ⅰ-7); 头部与卵黄囊分离, 卵黄囊体积减小; 眼内布满黑色素, 心脏搏动明显, 血液颜色加深呈淡红色; 口裂形成, 口与鳃盖有节律地闭合。
鳔雏形期:孵出后36h, 全长为6.7 mm, 肌节33对(图版Ⅰ-8); 卵黄囊体积进一步减小; 出现雏形鳔; 胸鳍呈扇型; 口裂加深; 听囊上缘开始出现黑色素, 之后色素增多, 在头背部变为似“)(”形状的色素纹, 同时吻端和眼睛后缘也出现少量黑色素, 之后随色素增多, 在头背部和两侧分别形成一对由吻端发出的分别到听囊上缘和眼睛后缘的两对射线, 卵黄囊上方近肌节处出现枝状色素带, 体背透明而无色素分布。
鳔一室期:孵出后54h, 全长为7.1 mm, 肌节33对(图版Ⅰ-9); 鳔初步形成, 未充气; 体背上缘肌节间出现一排纵向黑色素线, 且卵黄囊上方近肌节枝状色素带沿卵黄囊上缘延伸至肛门; 尾部动脉弧显红色; 肠道开始形成。
尾椎上翘期:孵出后72h, 全长为7.9 mm, 肌节33对(图版Ⅰ-10); 仅有少许卵黄囊剩余; 鳔充气, 呈椭圆形; 体背上缘肌节间纵向黑色素线加深, 与之对应的下缘肌节间开始出现一排纵向黑色素线; 卵黄囊上方与肌节间枝状色素带色素增多; 鳃盖后缘、15到17肌节及尾椎骨处体侧出现零星黑色素,以后黑色素进一步增多形成第一、第三、第六横斑雏形; 尾椎开始略微上翘; 背鳍褶前部稍隆起, 尾鳍褶开始分化, 其下叶开始出现放射状鳍条。
卵黄吸尽期:孵出后5d, 全长为9.5 mm, 肌节34对(图版Ⅰ-11a, b); 卵黄囊已吸尽; 口端位, 肠道贯通, 摄食量增加; 卵黄囊上方与肌节间枝状色素带色素增多; 第9到第11肌节、第19到第20肌节及第28到第29肌节处体测开始出现零星黑色素, 以后黑色素进一步增多形成第二、第四、第五横斑雏形; 体背不再透明, 皮肤开始变为淡黄色。
背鳍出现期:孵出后8d, 全长为12.8 mm, 肌节34对(图版Ⅰ-12a, b); 背鳍褶前部高高隆起, 形成背鳍原基; 尾鳍放射状鳍条增多, 达 13—15 根, 开始分叉; 消化道前端膨大, 形成食道, 口、肠道、肛门已连通。
臀鳍出现期:孵出后14d, 全长为14.5 mm, 肌节34对(图版Ⅰ-13a, b); 背鳍初步形成, 臀鳍开始出现。脊椎形成, 尾椎向上翘起, 尾鳍基本形成; 吻端开始长出须两对; 体背变为金黄色, 体表六条横斑雏形基本形成; 鳔分离成前后 2 室, 前室小、后室大。
稚鱼阶段稚鱼期:出膜20d, 全长达24.3 mm,各鳍条形成, 身体呈金黄色, 头部色素与体表六横纹色素完全形成, 出现鳞片与成鱼基本相似(图版Ⅰ-14a, b)。至此, 壮体沙鳅仔鱼发育过程完成, 进入稚鱼阶段。

图版Ⅰ 红水河来宾江段壮体沙鳅早期形态发育PlateⅠ Earlymorphogenesi of Botia robusta at Laibin section in Hongshuihe River 1. 尾牙期Tail bud; 2. 肌肉效应期Muscular contraction; 3. 心脏原基期Rudiment of heart; 4. 心脏搏动期Heart pulsation; 5. 胸鳍原基期Rudiment of pectoral fin; 6. 孵出期Hatching; 7. 眼黑色素期Melanoid eye; 8. 雏形鳔期Emergence of air bladder; 9. 鳔一室期One chamber air bladder; 10. 尾椎上翘期chordaltipliftin; 11. 卵黄吸尽期Exhaustion of York; 12 背鳍形成期Formation of dorsal fin; 13. 臀鳍形成期Formation of anal fin; 14. 稚鱼期Juvenile phase
2.2 红水河壮体沙鳅的产卵日期及其群体补充状况
采集的壮体沙鳅鱼卵主要处于尾牙期, 未采集到其鱼苗。根据鱼卵发育期推断, 红水河来宾江段壮体沙鳅产卵时间基本为4月30日至7月10日。采样期间, 共54d采集到壮体沙鳅的鱼卵, 总计1175颗,且有3批次较大规模的鱼卵流量, 分别为5月12—15日、6月17日及7月8日, 其中第1次的规模最大, 其中最大达203颗每网。 初步估算, 2017年4月中旬到8月底, 该江段壮体沙鳅总鱼卵补充群体量为2.23×108颗。所采集壮体沙鳅卵的产卵位置距采样点大约为15 km, 在上游的大湾镇附近。
2.3 鱼卵捕捞量与主要环境因子的关系
2017年4月16日至8月30日壮体沙鳅鱼卵单位捕捞努力量(CUPE)与流量及温度的关系如图 2、图 3所示。通过Pearson Correlation法, 对2017年4月中旬至8月底CUPE与水温、透明度、流量变化等因子进行相关性分析, 发现其单位捕捞努力量与水温显著相关(P<0.05) ; 而其单位捕捞努力量与透明度、流量变化相关性均不显著(P>0.05) 。
3 讨论
3.1 状体沙鳅早期发育形态特征
鳅科鱼类早期发育的研究资料相对较少, 目前珠江和长江水系分别有44种和66种鳅类, 仅有11种[3,12]有早期形态发育的研究资料。与前人的研究对比发现, 壮体沙鳅与花斑副沙鳅、长薄鳅的早期发育较为相似。为鉴别上的需要, 将三者早期发育的重要时期进行了归纳比较(表 1)。

图 2 红水河来宾段壮体沙鳅鱼卵单位捕捞努力量流量的关系Fig. 2 Relationship between the CUPE and runoff at Laibin section
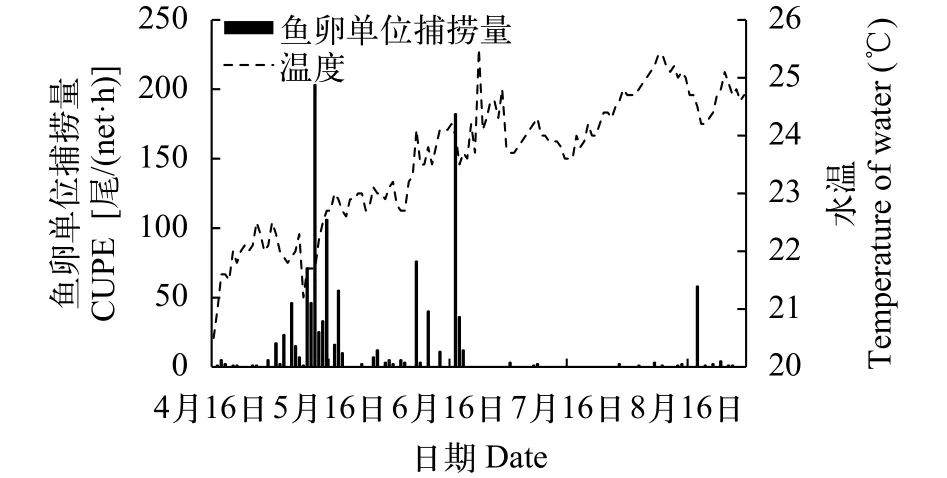
图 3 红水河来宾江段壮体沙鳅鱼卵单位捕捞量与水温关系Fig. 3 Relationship between the CUPE and water temperature at Laibin
从整体上看, 三者早期发育形态基本相同, 但壮体沙鳅胚后发育也具有不同于其他2种鱼类的特点, 主要表现在以下几个方面: 孵出时, 体色淡黄,肌节33对; 个体强壮, 全长约5.3 mm, 全长明显较花斑副沙鳅及长薄鳅长。鳔一室期, 头部色素明显,为两对由吻端发出的分别到听囊上缘和眼睛后缘的射线, 而长薄鳅头部色素为星状, 两者直观上容易判别; 该时期与花斑副沙鳅容易混淆, 但壮体沙鳅肌节仅为33对明显少于花斑副沙鳅的40对, 在显微镜下可明显区分。鳔二室期, 壮体沙鳅与花斑副沙鳅体横纹均基本形成, 但壮体沙鳅为6条, 而花斑副沙鳅为13条且其背鳍和尾鳍上亦有横纹; 长薄鳅体表则以星状色素为主。
3.2 西江来宾江段壮体沙鳅补充群体数量的资源前景预测
壮体沙鳅分布较广泛, 在干流或较大支流上游流水中产漂流性卵, 卵在流水中漂流发育, 后进入一定的水域继续发育。壮体沙鳅喜避光环境如(栖暗)漆暗的洞穴等, 同时对水体溶氧要求较高[13]。大坝修建会导致水文条件的剧变, 目前西江干流,从天峨县开始规划建设7座梯级, 已建6座[14]。随着来宾江段下游大藤峡水电站的建成蓄水, 该江段将逐渐变为库区, 水位抬高, 水深增加, 水面面积增加,水流变缓, 河流的水动力学过程发生较大的变化,库区鱼类组成将由“河流相”逐步向“湖泊相”演变。水文条件的一系列改变, 将导致水底洞穴、暗河压力增大, 溶氧降低, 导致壮体沙鳅的生存空间减少。已有研究认为, 水利工程的修建使圆口铜鱼的生存空间减少, 导致其不能完成生活史, 从而使其种群数量急剧减少[15,16], 同时圆口铜鱼为典型产漂流性卵的鱼类, 其鱼卵的比重略大于水的比重,当流速过缓时, 一部分鱼卵会沉入水底因缺氧而死亡[17]。故西江来宾段壮体沙鳅补充群体数量可能会受到大藤峡水电站的严重威胁。

表 1 壮体沙鳅与其他两种相同亚科鱼类胚后发育形态比较Tab. 1 Comparison of the postembryonic development of Botia robusta with two same species
