干旱胁迫对冬小麦水分关键期的生理特性和物质生产影响
2019-07-20张继波薛晓萍李鸿怡宋计平
张继波,薛晓萍,李 楠,李鸿怡,张 磊,宋计平
(1.山东省气候中心,山东 济南250031;2.临沂市气象局,山东 临沂276000;3.山东省农业科学院蔬菜与花卉研究所,山东 济南250031)
山东省是农业生产大省,2003 年以来,全省粮食总产实现了连续14 a 丰收,2016 年达到4.7×107吨,位列全国第三。在气候变暖的大背景下,受季风气候影响,山东省降水量年际变化大,年内分布及空间分布极不均匀,各地干旱灾害频发,并以春旱最为严重[1],对农业生产结构、种植制度和作物品种等产生重大影响[2,3]。开展水分关键期干旱胁迫对冬小麦生理特性及物质生产影响的研究,可以明确干旱胁迫对冬小麦生长发育及产量形成的影响机理,为保障冬小麦安全生产、科学防灾减灾提供技术支撑。
近年来,国内外关于干旱对作物生长发育及光合特性影响的研究已有相关报道[4-7]。土壤水分的变化势必引起作物生长发育和生理特性的响应,当作物遭受干旱胁迫时,作物体内含水量下降,气孔关闭,细胞膨压遭到破坏,导致生长延伸受到抑制[8,9]。干旱胁迫会严重影响作物光合作用、矿质元素代谢和次生代谢,导致活性氧积累,细胞膜损伤,叶绿素、蛋白质和脂类物质的降解,由此产生的丙二醛在某种程度上可作为判别细胞膜受损程度的指标[10-12]。抗氧化酶活性和种类随干旱胁迫的增强而增加,而增加的程度因作物的种类有所不同[13]。此外,干旱胁迫还会加速叶片衰老、抑制植株光合作用,减缓干物质积累及转运,最终造成作物减产[14-16]。作物任何发育期遭受干旱胁迫均会对其发育进程产生影响,其中,营养生长阶段水分亏缺会减缓叶片的生长速率并使得花期和生殖生长推迟,而生殖生长阶段水分亏缺则会加速作物的成熟与衰老[17-19]。
农业生态系统中,干旱可能发生在作物的任何生长发育阶段,而拔节期—孕穗期是冬小麦由冬前营养生长转变为营养与生殖生长同时并进的时期,是决定冬小麦最终产量的关键时期[18]。本研究针对农业生产的实际,结合前人的研究成果,在临沂设施农业气象试验站水分控制场设计冬小麦水分关键期水分控制试验,模拟不同水分处理下土壤水分变化规律,系统研究水分关键期冬小麦光合生理特性、抗氧化酶活性及产量结构对干旱胁迫的响应,以期为科学评估春季干旱对冬小麦生长发育及产量形成的影响提供依据。
1 材料与方法
1.1 试验设计
试验于2016—2017 年和2017—2018 年冬小麦生长季内在临沂设施农业气象试验站自动控制遮雨棚内的水分控制场进行。水分控制场位于118°14′54″E、35°28′30″N,包括15 个试验小区,南侧4 m 处为3 个雨养对照小区,小区内土壤均为砂壤,小区面积为16 m2(4 m×4 m)、深2 m,小区间距0.2 m,四周及底部水泥墙均做防水防渗处理,水分控制场地上部分安装自动移动式遮雨棚,遮雨棚长23 m,宽14 m,高5 m,雨雪时将遮雨棚关闭棚遮蔽试验小区,其余时段为自然光照,遮雨棚顶部及四周为透光率85%的阳光板,可保证雨雪天气时作物最大限度接受散射光。供试材料为冬小麦品种“齐麦2 号”,2016—2017 年和2017—2018 年播期均为10 月8 日,播种方式为条播,密度为每平方米1000 株,播种后供水充分,保证出苗,除水分控制期间外其他时段均保证所有处理处于水肥条件适宜且一致。
在冬小麦水分关键期(拔节期~扬花期)设置5个梯度水分处理及1 个雨养对照,分别标记为T1、T2、T3、T4、T5 和CK,重复3 次,共18 个试验小区,具体设计见图1。冬小麦返青后隔天观测发育期,逢8 进行人工测墒,进入拔节期前,将各处理20 cm 土壤相对湿度控制在60%左右,进入拔节期后参照临沂地区冬小麦水分关键期(4 月1—30 日)常年降水量(约30 mm)设置T1、T2、T3、T4、T5 共5 个水分处理。其中,水分控制期间T1 处理维持在适宜水平(20 cm 土壤相对湿度80%左右),T2、T3、T4、T5 处理分别较水分关键期常年降水量减少20%、50%、75%、100%进行一次性补水,直至扬花期不再补水,扬花期后复水直至冬小麦成熟。
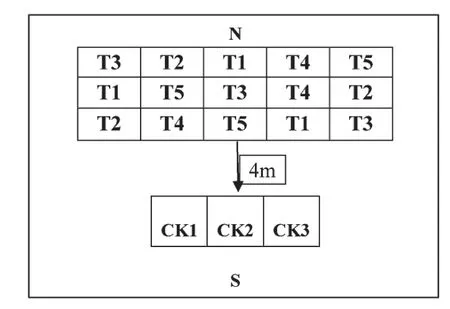
图1 水分控制场试验小区分布
不同水分处理下20 cm 土壤相对湿度变化情况见图2。由图2 可知,4 月1 日冬小麦进入拔节期,除CK 和T5 外,T1、T2、T3、T4 处理20 cm 土壤相对湿度均明显上升,其中,T1、T2、T3 处理超过80%,T4处理超过70%;水分控制期间,气温回升,作物蒸腾和田间蒸散增强,除CK 和T1 外,T2、T3、T4、T5 处理20 cm 土壤相对湿度均显著下降。根据土壤水分动态监测干旱判别标准(表1),冬小麦扬花期时,T2、T3 处理达到轻度干旱,T4、T5 处理达到重度干旱。
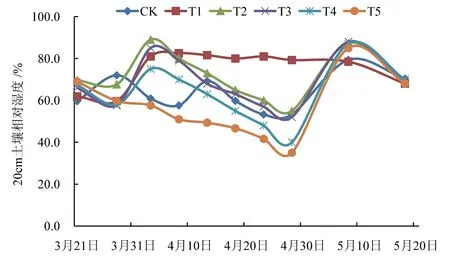
图2 不同水分处理20 cm 土壤相对湿度变化

表1 土壤水分动态监测干旱判别标准
1.2 测定项目与方法
1.2.1 叶片光响应曲线测定
利用Li-6400 便携式光合作用测定系统(Licor,USA),在晴天9:00—12:00 采用Li-6400-02B红蓝光叶室测定Pn-PAR 光响应曲线,各处理选择功能叶片测定2 株。水分控制期间测定间隔为10 d,拔节期前、灌浆期和乳熟期择期分别测定1 次。
通过开放式气路,设定温度为25 ℃,大气CO2浓度(Ca)为400 μmol/mol,设定光强梯度为1800、1600、1400、1200、1000、800、600、400、350、300、200、150、100、50、20、0 μmol··。测定叶片在每一光强下的净光合速率(,μmol··)、气孔导度(Gs,mmol··)、胞间浓度(Ci,μmol/mol)等光合参数,计算气孔限制值(Ls)=1-Ci/Ca。
用直角双曲线模型在SPSS15.0 中对各处理的光响应曲线进行拟合,得到光饱和时的最大光合速率()、表观量子效率(),直角双曲线模型[20]表达式如下:
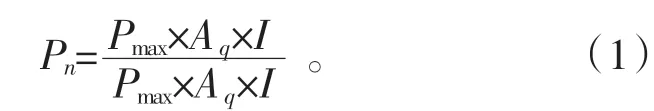
1.2.2 保护酶活性及丙二醛(MDA)含量测定
水分控制期间,在9:00—10:00 选取长势均匀植株顶端以下的第5~8 位叶片,采摘后用液氮速冻,然后保存于-40 ℃超低温冰箱,在山东省农业科学院蔬菜与花卉研究所实验室内完成各项酶活性的测定,取样频次与光响应曲线测定同步。采用氮蓝四唑法测定超氧化物歧化酶(SOD)活性,采用愈创木酚显色法测定过氧化物酶(POD)活性,采用紫外分光光度计测定过氧化氢酶(CAT)活性,采用硫代巴比妥酸法测定MDA 含量[21]。
1.2.3 灌浆速率测定
冬小麦进入灌浆期后,各处理分别选取1 m 样段进行麦穗取样,计算麦穗数量,并测定鲜重、干重,之后间隔10 d、20 d 分别测定一次,计算得到灌浆速率。
1.2.4 产量结构测定
冬小麦成熟后,各处理均匀选取样株进行收获,晾晒后室内考种,考种项目包括穗粒数、千粒重等,测定其理论产量。
1.3 数据处理
试验数据采用Microsoft Excel 2003 进行整理,SAS 9.0 进行方差分析,采用邓肯新复极级差法分析不同处理间显著差异。
2 结果与分析
2.1 干旱胁迫对冬小麦叶片光合特性的影响
水分控制期间,除T1 外,其他各处理土壤相对湿度不同程度降低,干旱胁迫逐渐增强,叶片气孔关闭以减少蒸腾作用造成的水分散失,进而阻碍CO2进入叶片,对冬小麦叶片光合作用产生影响。水分控制期间冬小麦叶片光合生理参数变化显著(图3)。由图3 可知,饱和光强下,冬小麦叶片、Gs、Ci 均随干旱胁迫的加剧呈降低趋势,干旱胁迫持续时间越长,、Gs 降低幅度越大,水分处理28 d 后,T2、T3、T4、T5 处 理、Gs 分 别 较T1 降 低76.6%~90.0%和70.8%~96.7%,均达到显著水平,T5 处理Pmax、Gs 降低幅度均最大。T2、T3 处理冬小麦叶片Pmax、Gs、Ci 均随干旱胁迫的加剧而降低,Ls 升高,Ci和Ls 变化趋势相反,气孔因素是轻度干旱胁迫下冬小麦叶片光合速率降低的主要原因;饱和光强下,Ls随干旱胁迫的加剧呈升高趋势,水分处理13 d 后,T4、T5 处理Ci 升高,Ls 降低,重度干旱胁迫下导致冬小麦叶片光合速率降低的主要原因是非气孔因素。
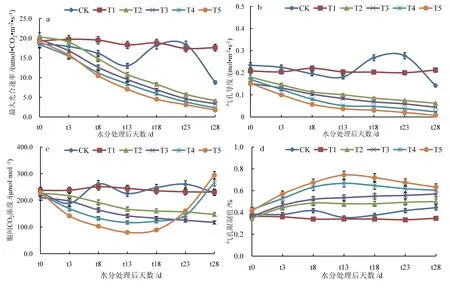
图3 不同水分处理对冬小麦叶片光合生理参数的影响
扬花期冬小麦叶片光响应曲线对干旱胁迫的响应见图4。由图4 可知,不同水分处理下冬小麦叶片光合速率随光照强度的增强均呈上升趋势,并在光饱和点达到最大值,随着光照强度进一步增强,T2、T3、T4、T5 处理冬小麦叶片光合速率均出现不同程度降低,T5 处理降低幅度最大,降幅达87.6%,T4 处理次之,T2 处理最小,降幅为36.7%。干旱胁迫下冬小麦叶片出现“光抑制”现象,且随干旱胁迫的加剧而加重。
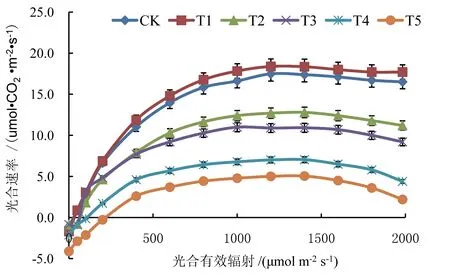
图4 不同水分处理对扬花期冬小麦叶片光响应曲线的影响
2.2 干旱胁迫对冬小麦叶片抗氧化酶和丙二醛含量的影响
不同水分处理下SOD 酶、POD 酶、CAT 酶活性及MDA 含量的变化情况见图5。由图5 可知,T2、T3处理冬小麦叶片SOD 酶、POD 酶、CAT 酶活性均呈上升趋势,均在水分处理后15 d 达到最大,分别为410.0 U·g-1·min-1、395.0 U·g-1·min-1、18.6 U·g-1·min-1和463.0 U·g-1·min-1、451.0 U·g-1·min-1、19.9 U·g-1·min-1,复水后能够恢复至正常水平;随着干旱胁迫的加剧,T4、T5 处理冬小麦叶片SOD 酶、POD 酶、CAT酶活性则呈先上升后降低的趋势,同样在水分处理第15 日达到最大,分别为547.0 U·g-1·min-1、520.0 U·g-1·min-1、22.0 U·g-1·min-1和616.0 U·g-1·min-1、572.0 U·g-1·min-1、23.4 U·g-1·min-1,均显著高于T2、T3 处理,复水后缓慢降低,无法恢复至正常水平。干旱胁迫下,T2、T3、T4、T5 处理MDA 含量均呈上升趋势,复水后T2、T3 处理MDA 含量基本可以恢复至正常水平,而T4、T5 处理无法恢复至正常水平,重度干旱胁迫导致冬小麦叶片抗氧化酶系统遭受不可逆损伤。
2.3 干旱胁迫对冬小麦灌浆速率和产量结构的影响
随着干旱胁迫的加剧,冬小麦灌浆速率显著降低(图6)。T1 处理和CK 由于前期水分充足,冬小麦灌浆充分,灌浆速率分别为0.049 g/d 和0.041 g/d,单穗干物质重差异不显著;T2、T3 处理灌浆速率分别为0.038 g/d 和0.032 g/d,单穗干物质重分别较T1 减少40.4%和20.7%,T4、T5 处理灌浆极不充分,单穗干物质重分别较T1 减少64.1%和70.0%,这可能由于干旱胁迫导致冬小麦无法正常授粉造成的。
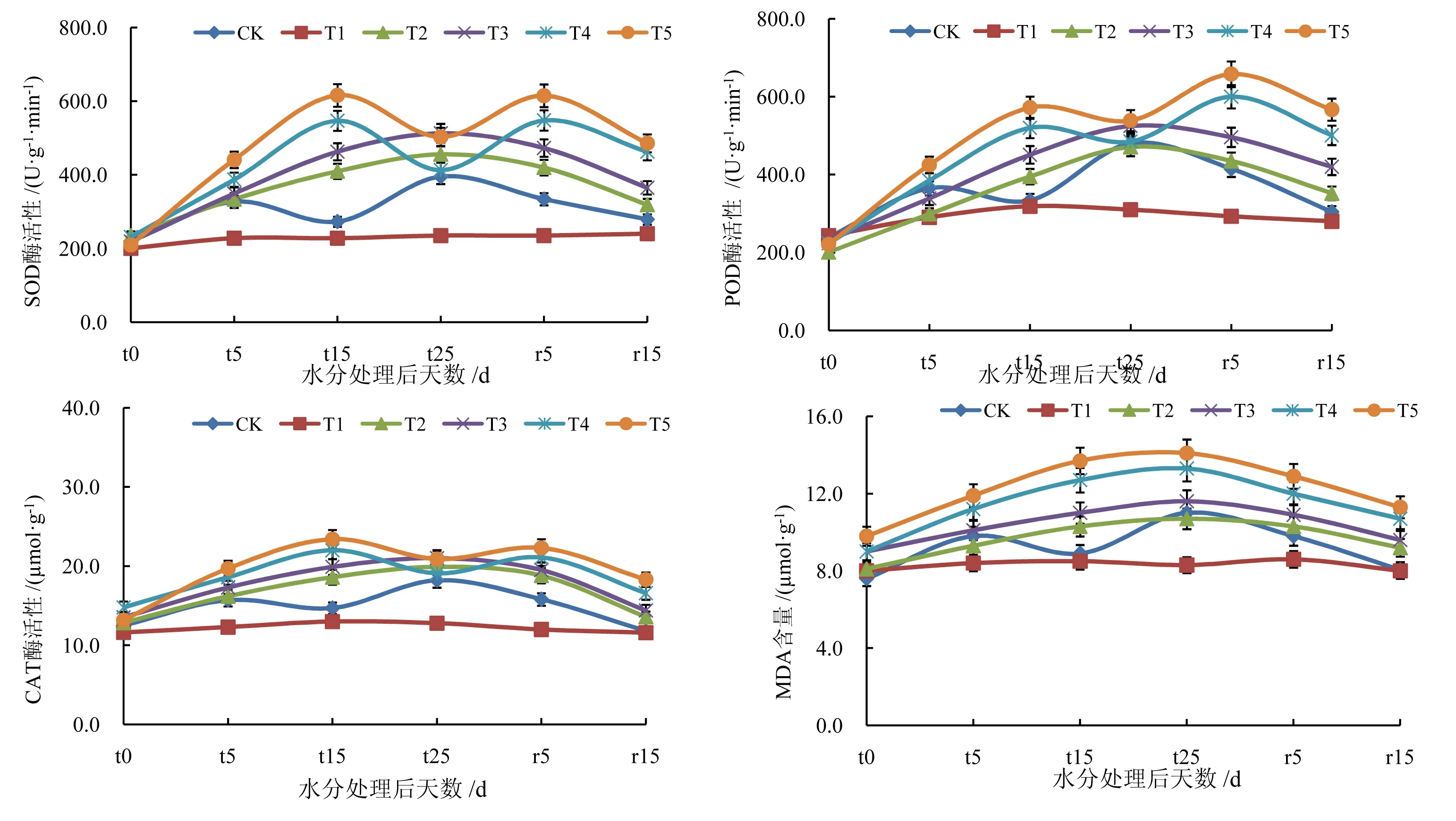
图5 不同水分处理对冬小麦叶片抗氧化酶和丙二醛含量的影响

图6 不同水分处理对冬小麦灌浆速率的影响
随干旱胁迫的加剧,冬小麦穗粒数、千粒重、理论产量均呈下降趋势,而不孕小穗率则呈升高趋势(图7)。由图7 可知,T1 处理冬小麦不孕小穗率仅为2.0%,CK 与T1 几乎无差异,T2、T3 处理和T4、T5 处理不孕小穗率分别超过10.0%和30.0%,均显著高于T1 处理。T1 处理和CK 穗粒数分别为43.2个和45.3 个,差异不显著,而T2、T3、T4、T5 处理穗粒数则均显著低于T1 处理,分别较T1 处理降低17.1%、30.3%、51.6%和56.7%,其中,T4、T5 处理间差异不显著。由于灌浆不充分,T4、T5 处理千粒重均显著小于T1 处理,分别较T1 处理降低10.6%和11.8%,而T2、T3 处理千粒重则与T1 处理相差不大。干旱胁迫对冬小麦理论产量造成不可逆的影响,T2、T3、T4、T5 处理理论产量均显著低于T1 处理,T1 处理理论产量为8 326.5 kg/hm2,而T2、T3 处理为6 228.0 kg/hm2和5 418.0 kg/hm2,分别较T1 减少25.2%和34.9%,T4、T5 处理为4 264.5 kg/hm2和3 616.5 kg/hm2, 分 别 较T1 处 理 减 少48.8%和56.6%。
3 结论
(1)干旱胁迫显著降低冬小麦叶片光合速率,轻度干旱胁迫和重度干旱胁迫下冬小麦叶片光合速率的降低分别是由气孔因素和非气孔因素造成的。干旱胁迫会造成冬小麦出现“光抑制”现象,干旱胁迫越重,“光抑制”越重。
(2)干旱胁迫下冬小麦叶片抗氧化酶系统的紊乱是造成叶片光合速率降低的内在因素,随干旱胁迫的加剧,冬小麦叶片SOD、POD 和CAT 酶活性先升高后降低,当干旱胁迫达到重度时,SOD、POD 和CAT 酶活性受到抑制,MDA 含量显著升高,对冬小麦叶片膜系统造成不可逆损伤,冬小麦叶片光合能力显著降低。
(3)水分关键期干旱胁迫造成冬小麦灌浆速率、穗粒数、千粒重、理论产量均显著降低,且干旱胁迫越重,降低幅度越大;而不孕小穗率则呈升高趋势。
4 讨论
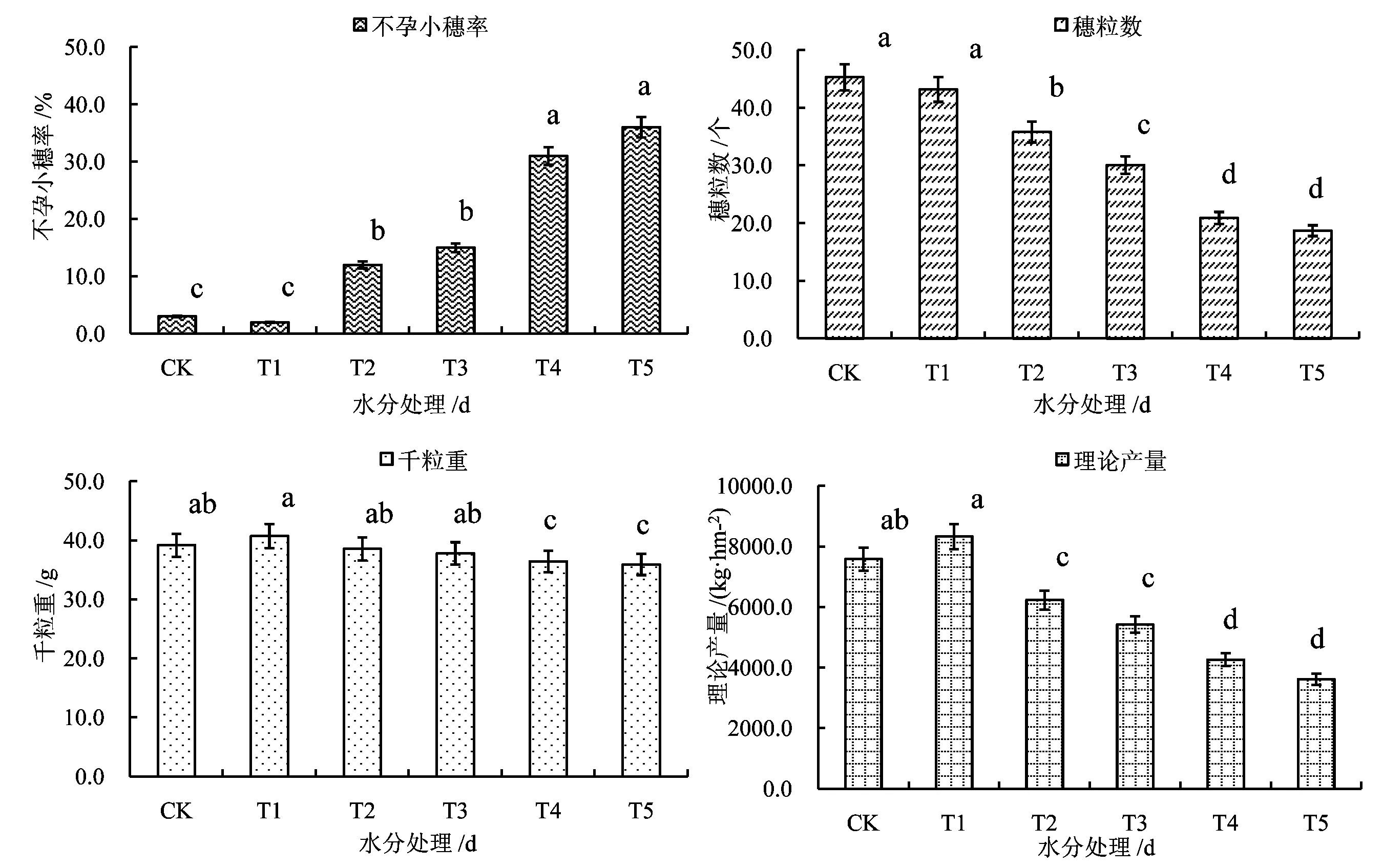
图7 不同水分处理对冬小麦产量结构的影响
光合作用是作物干物质积累和产量形成的基础[22]。干旱胁迫对作物叶片光合参数的影响与叶片气孔的开放程度有关,作物遭受干旱胁迫时,首先通过关闭叶片气孔以减少蒸腾作用,进而阻止CO2进入叶片,从而对叶片光合作用产生影响。干旱胁迫下冬小麦叶片光合速率显著降低,且随干旱胁迫的加剧降低幅度增大,而光合速率降低的原因却有所不同,干旱胁迫下气孔因素和非气孔因素是造成冬小麦叶片光合速率降低的主要原因,Ci 和Ls 对干旱胁迫的响应是判断造成冬小麦叶片光合速率降低原因的主要依据[14,21,23]。
干旱胁迫下,冬小麦叶片PSⅡ的活性降低、活性氧积累,造成冬小麦叶片细胞膜脂过氧化程度升高,进而对冬小麦代谢过程产生影响。本研究表明:干旱胁迫会诱导冬小麦叶片SOD 酶、POD 酶和CAT 酶活性升高,进而降低MDA 含量,减轻干旱胁迫对冬小麦叶片PSⅡ的影响;随着干旱胁迫的加剧,重度干旱胁迫下冬小麦叶片SOD 酶、POD 酶和CAT 酶活性受到抑制, 无法清除叶片内过多的MDA,导致冬小麦叶片膜系统遭受不可逆的破坏,冬小麦叶片光合能力显著降低,这与张仁和等[10]的研究结论基本一致。
干旱胁迫是限制冬小麦产量提高的主要因素之一[24]。干旱胁迫下冬小麦授粉不充分,灌浆期缩短,灌浆速率、千粒重显著降低,不孕小穗率升高,进而导致冬小麦减产。本研究表明:干旱胁迫下T2、T3处理冬小麦灌浆速率分别较T1 处理降低22.4%、34.7%,而T4、T5 处理较T1 处理降低超过50%,严重制约了冬小麦的产量提高。理论产量是预估冬小麦实际产量的有效手段,T2、T3、T4、T5 处理冬小麦理论产量仅为T1 处理的74.8%、65.1%、51.2%和43.4%,随干旱胁迫的加剧,冬小麦产量降低越显著,这可能是由于干旱胁迫下植株体内激素平衡遭到破坏造成的[24-25]。
通过研究水分关键期干旱胁迫对冬小麦叶片光合生理、抗氧化酶活性及产量结构的影响,探讨干旱胁迫对冬小麦生长发育及产量形成的影响机理,为明确干旱胁迫下冬小麦的致灾机理、灾损评估及应灾减灾提供依据,今后仍应进一步开展不同发育期干旱胁迫对冬小麦生长发育及产量结构影响的研究。
