高产纤维素酶里氏木霉菌株诱变选育及其发酵条件优化
2019-07-20章亭洲王腾浩
葛 青,章亭洲,2*,王腾浩,赵 艳
(1.浙江科峰生物科技有限公司,浙江 海宁 314400;2.浙江工商大学,杭州 310000)
纤维素酶可以分解纤维素,添加纤维素酶能够改善粗饲料的柔韧性,并破坏饲料中粗纤维等抗营养物质,从而提高动物采食量,改善动物消化道健康[1]。添加纤维素酶还可以补充动物体内内源酶的不足,提高对粗纤维的利用率,同时还可以改善消化道酶系组成、酶量及活性,从而提高对营养物质的利用率[2]。因此,纤维素酶对于现代饲料产业具有重要的现实意义和应用价值,但是目前饲用纤维素酶制剂仍然存在功能单一、稳定性差、不耐热、酶活性低、产率低且成本高等缺陷[3-4]。这是影响纤维素酶制剂广泛应用于饲料中的瓶颈问题,筛选高产纤维素酶生产菌株具有重要意义[5-6]。
里氏木霉是一种好氧的丝状真菌,其分泌的纤维素酶是胞外酶,经初提和分离纯化就可得到纤维素酶制剂[7-8]。里氏木霉产纤维素酶量高、稳定性好、适应性强,且可以通过物理和化学诱变获取高产菌株,便于形成规模化生产和管理[9]。研究采用里氏木霉的诱变方法,改造菌株同时优化产酶培养基及发酵条件,并采用培养条件易于控制、生产效率高且不易染菌的液态发酵方法培养菌株,具有重要的现实意义和应用价值。因此,本研究利用常压室温等离子体(ARTP)诱变法处理里氏木霉菌株,选育纤维素酶高产突变株,优化高产纤维素酶突变菌株发酵工艺,确定最适发酵条件,为高产纤维素酶菌株的开发和应用提供依据。
1 材料与方法
1.1 试验材料
1.1.1 菌种
里氏木霉RUT-C30 购自广东微生物菌种保藏中心。
1.1.2 培养基
微晶纤维素刚果红培养基:羧甲基纤维素钠10 g,乳糖3 g,硫酸铵2 g,磷酸氢二钾2.0 g,七水合硫酸镁0.5 g,刚果红0.2 g,琼脂粉20 g,用蒸馏水定容至1 L。
种子培养基:葡萄糖15 g,酵母浸膏20 g,硫酸镁0.8 g,磷酸氢二钾6 g,氯化钙1.0 g,硫酸铵2.5 g,Mandels 微量元素营养盐1 mL,吐温-80 2 mL,pH 4.8,用蒸馏水定容至1 L。
发酵产酶培养基:乳糖18 g,玉米桨粉12 g,微晶纤维素10 g,硫酸镁1 g,硫酸铵0.5 g,氯化钙0.5 g,Mandels 微量元素营养盐1 mL,吐温-80 2 mL,pH 4.8,用蒸馏水定容至1 L。
1.2 诱变和筛选
1.2.1 孢子悬液制备
取活化培养的里氏木霉平板,用无菌水冲洗,接入种子培养基,30 ℃,150 r·min-1培养。在此过程中记录孢子萌动过程,以萌动率20%~30%为最佳萌动时间。
1.2.2 ARTP诱变
将里氏木霉孢子液用无菌水稀释至107~108个·mL-1进行常压室温等离子体(ARTP)诱变。照射距离为2 mm,诱变时间分别为0、40、60、80、100、120、140、160、180、200、220、240、260 s。吸取不同诱变时间的菌液均匀涂布于种子培养基中,30 ℃恒温恒湿培养48 h,记录菌落数,以无菌水处理的作为空白对照,计算致死率,公式见式1。
致死率/%=(处理菌落数-对照菌落数)/对照菌落数×100% (1)
1.2.3 最优突变菌筛选
将诱变得到的里氏木霉涂布于微晶纤维素刚果红培养基上,以原菌里氏木霉在微晶纤维素刚果红培养基上产生的透明圈菌落大小为对照,挑选出平板上突变菌长得快、透明圈大、菌丝短、产孢子晚的单菌落。用0.9%的生理盐水清洗,然后涂到传代培养基上,培养4~6 d,接种至固体发酵培养基上进行发酵,测定其总酶活(FPU),比较其酶活性大小以确定高产纤维素酶分解菌株。
1.3 酶活力测定
以滤纸作为底物测定纤维素酶总酶活:将滤纸条置于装有1 mL 柠檬酸缓冲液(50 mmol·L-1,pH 4.8)的试管中作为反应底物,然后加入500 μL经稀释的发酵上清液,50 ℃恒温振荡反应1 h,反应结束后立即加入3,5-二硝基水杨酸溶液3 mL,沸水浴5 min,迅速冷却至室温,加入去离子水20 mL,混合均匀后测定540 nm处的吸光值[11]。空白对照为柠檬酸缓冲液(50 mmol·L-1,pH 4.8)500 μL。
酶活的表示用U·mL-1上清液来衡量。其中一个酶活单位(U)定义为指在1 min 内释放1 μmol 底物所需要的酶量。
1.4 突变菌产酶发酵条件优化
1.4.1 单因素实验
以基础培养基为基准,30 ℃恒温培养,依次对发酵时间(12、24、36、48、60、66、72、78、84、90、96、102、108、114、120、132、144 h)、硫酸铵浓度(0、0.5、1、1.5、2、2.5 g·L-1)、微晶纤维素浓度(6、8、10、12、14 g·L-1)、接种量(7%、8%、9%、10%、11%、12%、13%)及搅拌速度(200、300、400、500、600 r·min-1)进行单因素实验,保持其他条件恒定(乳糖10 g·L-1,微晶纤维素10 g·L-1,玉米浆粉12 g·L-1,(NH4)2SO41.5 g·L-1,MgSO41.2 g·L-1,CaCl21.4 g·L-1,Mandels 微量元素营养盐1 mL·L-1,吐温-80 2 mL·L-1,pH 4.8,搅拌转速为400 r·min-1,通气量2 vvm。每个重复3次。测定不同发酵条件下突变菌里氏木霉滤纸酶活性,确定这些因素对突变菌里氏木霉产酶的影响,并得到最佳产酶条件。
1.4.2 正交实验
根据单因素实验结果,选择单因素实验中对产酶影响较大的4 个因素,每个因素设3 个水平,即采用L9(34)正交实验,对里氏木霉突变菌的产滤纸酶的最优固体发酵条件组合进行优化。
2 结果与分析
2.1 突变株筛选
2.1.1 ARTP诱变与突变菌筛选
将里氏木霉孢子萌发后进行ARTP 诱变,最佳照射时间为200~240 s(致死率达到98%)。经多轮ARTP 诱变,挑选出各轮诱变菌透明圈直径/菌落直径较大的初筛突变菌。初筛突变菌经摇瓶发酵进行复筛,突变菌里氏木霉的产酶能力见表1。
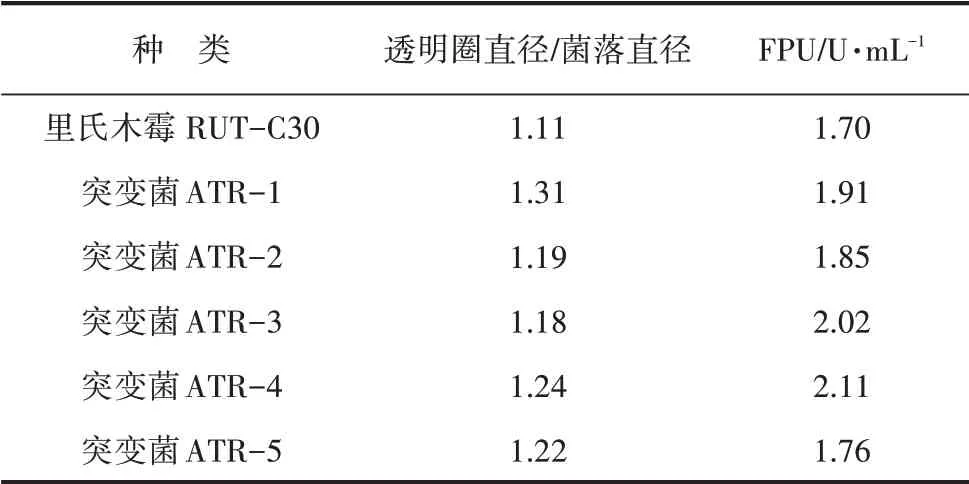
表1 里氏木霉RUT-C30 突变菌产酶能力
由表1可知,通过突变菌菌落大小比较,获得5 个突变菌,经摇瓶发酵复筛发现,突变菌里氏木霉ATR-4的滤纸酶活性较原菌里氏木霉有所提高。鉴于里氏木霉ATR-4 滤纸酶活(FPU)最高,可达2.01 U·mL-1,因此选择KFDY-4菌株作为产酶发酵条件优化实验的突变菌。
2.2 突变菌里氏木霉ATR-4产酶发酵条件优化
2.2.1 发酵时间对里氏木霉ATR-4产酶能力的影响
发酵时间对里氏木霉ATR-4 产酶能力的影响见图1。

图1 发酵时间对里氏木霉ATR-4产酶能力的影响
由图1可知,发酵时间对里氏木霉ATR-4产酶能力的影响较大。随着发酵时间的延长,滤纸酶活性呈现先升高再降低再升高的变化趋势,发酵时间84 h 时滤纸酶活达到最高,随后滤纸酶活性随着发酵时间的延长而呈下降趋势。由于发酵时间越短,对降低生产成本较为有利,因此选择里氏木霉ATR-4最佳发酵时间定为84 h。
2.2.2 硫酸铵浓度对里氏木霉ATR-4 产酶能力的影响
硫酸铵浓度对里氏木霉ATR-4 产酶能力的影响见图2。
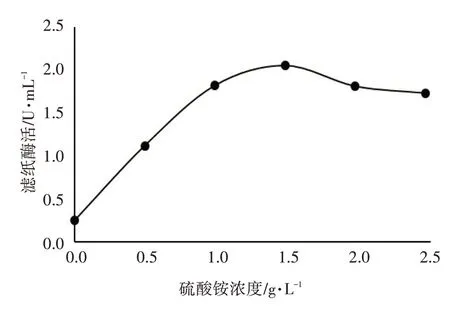
图2 硫酸铵浓度对里氏木霉ATR-4产酶能力的影响
由图2 可知,滤纸酶活性在硫酸铵浓度为1.5 g·L-1时最高,显著高于硫酸铵浓度为0、0.5、2.5 g·L-1时(P<0.05)。因此,选择里氏木霉ATR-4 最佳的硫酸铵浓度定为1.5 g·L-1。
2.2.3 微晶纤维素浓度对里氏木霉ATR-4 产酶能力的影响
微晶纤维素浓度对里氏木霉ATR-4 产酶能力的影响见图3。
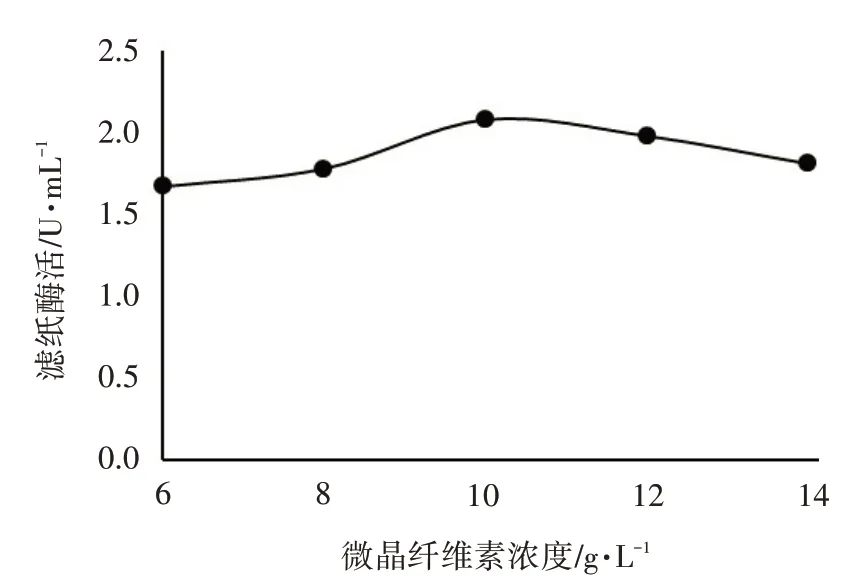
图3 微晶纤维素浓度对里氏木霉ATR-4产酶能力的影响
由图3 可知,微晶纤维素对滤纸酶活性无显著影响,微晶纤维素浓度在10 g·L-1时,滤纸酶活性相对较高。综合考虑成本,里氏木霉ATR-4 产酶的最佳微晶纤维素浓度定为10 g·L-1。
2.2.4 接种量对里氏木霉ATR-4产酶能力的影响
接种量对里氏木霉ATR-4 产酶能力的影响见图4。
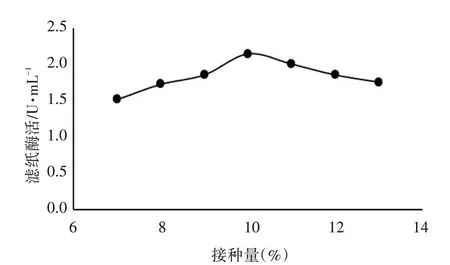
图4 接种量对里氏木霉ATR-4产酶能力的影响
由图4 可知,接种量在7%~10%时,滤纸酶活性随着接种量的增加而提高,接种量>10%时,滤纸酶活性随着接种量的增加而下降,当接种量为10%时,滤纸酶活性达最高,为2.15 U·mL-1。因此,里氏木霉ATR-4产酶的最佳接种量定为10%。
2.2.5 搅拌速度对里氏木霉ATR-4 产酶能力的影响
搅拌速度对里氏木霉ATR-4 产酶能力的影响见图5。
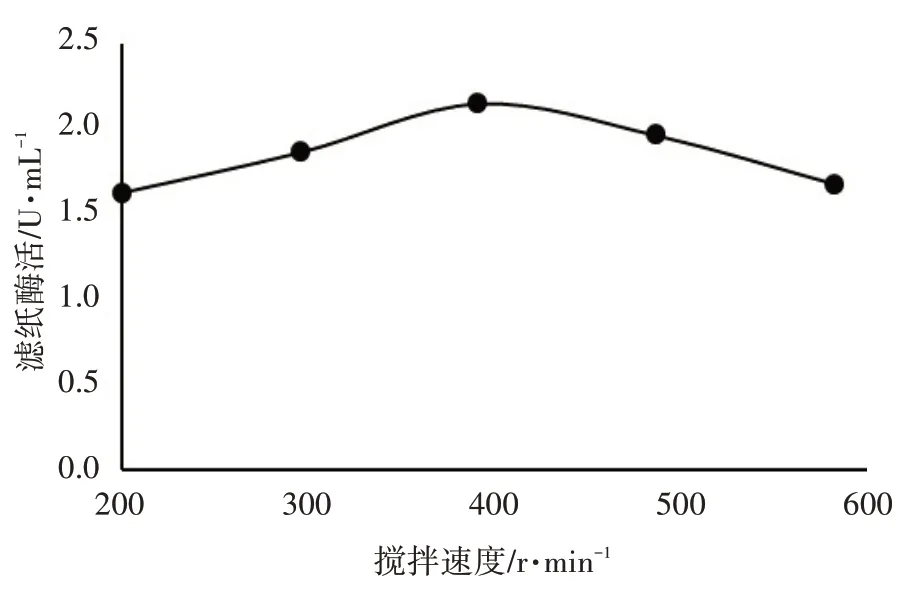
图5 搅拌速度对里氏木霉ATR-4产酶能力的影响
由图5 可知,搅拌速度在400 r·min-1时,滤纸酶活性最高,显著高于搅拌速度200、600 r·min-1时(P<0.05)。因此,里氏木霉ATR-4 产酶的最佳搅拌速度定为400 r·min-1。
2.2.6 正交实验
通过单因素实验初步确定里氏木霉ATR-4 产酶的最佳工艺条件为:发酵时间84 h,硫酸铵浓度1.5 g·L-1,微晶纤维素浓度10 g·L-1,接种量10%,搅拌速度400 r·min-1。根据单因素实验的结果,选择对里氏木霉ATR-4 固体发酵产酶能力影响较大的4 个因素(发酵时间、硫酸铵浓度、接种量、搅拌速度)进行L9(34)的正交实验,每个试验进行3次。正交实验因素水平表见表2。
正交实验结果及极差分析见表3,以极差大小确定因素主次顺序。从极差分析可知,影响里氏木霉ATR-4 产酶的主要因素是搅拌速度,其次是发酵时间和接种量,硫酸铵浓度的影响最小。里氏木霉ATR-4产酶的最佳组合为A1B1C2D3,即发酵时间78 h,硫酸铵浓度1 g·L-1,接种量10%,搅拌速度400 r·min-1。

表2 正交实验因数水平表L 9(34)

表3 正交实验结果
2.2.7 正交实验验证
在上述正交实验优化条件下,对里氏木霉ATR-4 发酵产酶活性的最适发酵条件进行验证,平行实验3次,测得滤纸酶活性为4.57 U·mL-1,该结果大于单因素实验最佳条件的得率。说明各因素间相互作用可以提高里氏木霉ATR-4 产滤纸酶活性,达到更好的提取效果。
3 结 论
采用常压室温等离子体(ARTP)诱变技术处理里氏木霉Trichoderma reesei RUT-C30菌株,经液体发酵筛选,选择纤维素酶活性较高菌株并对其液体发酵条件进行优化。在诱变时间240 s 条件下获得11 株突变里氏木霉ATR-4,其滤纸酶活(FPU)最高可达2.01 U·mL-1。对突变里氏木霉菌株ATR-4的发酵条件优化,筛选得到最佳产酶培养条件为:发酵时间78 h,硫酸铵浓度1 g·L-1,接种量10%,搅拌速度400 r·min-1,乳糖10 g·L-1,微晶纤维素10 g·L-1,玉米浆粉12 g·L-1,MgSO41.2 g·L-1,CaCl21.4 g·L-1,Mandels 微量元素营养盐1 mL·L-1,吐温80 2 mL·L-1,pH 4.8,发酵温度30 ℃,通气量2 vvm。在此条件下进行验证实验,最高酶活可达4.57 U·mL-1。本研究结果表明,常压室温等离子体(ARTP)诱变可有效对里氏木霉进行诱变育种,改善其产酶能力。研究选育获得产纤维素酶能力较稳定的突变里氏木霉菌株ATR-4,可为后续大规模培养与应用提供重要依据。
