Sporamin suppresses growth of xenografted colorectal carcinoma in athymic BALB/c mice by inhibiting liver β-catenin and vascular endothelial growth factor expression
2019-07-12ChunYangJingJieZhangXiaoPengZhangRongXiaoPengGaoLi
Chun Yang, Jing-Jie Zhang, Xiao-Peng Zhang, Rong Xiao, Peng-Gao Li
Abstract BACKGROUND Colorectal cancer (CRC) is the third most common malignancy of the digestive tract and the fifth leading cause of cancer-related mortality in China. Sporamin, a Kunitz-type trypsin inhibitor isolated from sweet potato, is a potential anti-cancer agent with activities against a number of malignant tumor cells in vitro. The liver secretes a myriad of endocrine factors that may facilitate the growth and transformation of tumors in the development of CRC.AIM To investigate the effects of sporamin on liver morphology and biomarkers of xenografted CRC in the liver of athymic BALB/c mice.METHODS Twenty-seven male BALB/c nude mice were randomly divided into control,vehicle, and sporamin groups. Mice in the latter two groups were intraperitoneally xenografted with LoVo colorectal carcinoma cells and intragastrically infused with saline or sporamin (0.5 g/kg body weight/d),respectively, for 3 wk. Hematoxylin and eosin (HE) staining of the sections was performed to observe morphological changes in hepatic tissue and real-time fluorescent quantitative PCR (qPCR) and enzyme-linked immunosorbent assay(ELISA) were used to measure the expression of β-catenin and vascular endothelial growth factor (VEGF) in the liver.RESULTS Sporamin significantly reduced the number and weight of tumor nodules formed in the abdominal cavity. Compared with the vehicle group, the mean tumor weight (± SD) in the sporamin group was significantly reduced (0.44 ± 0.10 g vs accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0)license, which permits others to distribute, remix, adapt, build upon this work non-commercially,and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See:http://creativecommons.org/licen ses/by-nc/4.0/Manuscript source: Unsolicited manuscript Received: February 23, 2019 Peer-review started: February 23,2019 First decision: April 4, 2019 Revised: May 14, 2019 Accepted: May 31, 2019 Article in press: June 1, 2019 Published online: July 7, 2019 P-Reviewer: Demir Y, Mohamed SY S-Editor: Ma RY L-Editor: Wang TQ E-Editor: Ma YJ 0.26 ± 0.15 g) and the total number of tumors decreased from 93 to 55. HE staining showed that enlargement of the nucleus and synthesis of proteins within hepatocytes, as well as infiltration of inflammatory cells into the liver, were attenuated by sporamin. Immunohistochemical staining and ELISA showed that the concentrations of β-catenin and VEGF in the liver were significantly reduced by sporamin. Compared with the vehicle group, the expression of β-catenin measured in integrated optical density units per area was reduced in the sporamin group (47.29 ± 9.10 vs 26.14 ± 1.72; P = 0.003). Expression of VEGF was also reduced after sporamin intervention from 20.78 ± 2.06 in the vehicle group to 15.80 ± 1.09 in the sporamin group (P = 0.021). Compared with the vehicle group,the concentration of β-catenin decreased from 134.42 ± 22.04 pg/mL to 109.07 ±9.65 pg/mL after sporamin intervention (P = 0.00002). qPCR indicated that compared to the vehicle group, relative mRNA expression of β-catenin and VEGF in the liver of mice in the sporamin-treated group was significantly reduced to 71% ± 1% (P = 0.000001) and 23% ± 7% (P = 0.00002), respectively, of the vehicle group levels.
Key words: Sporamin; Colorectal cancer; Liver; Vascular endothelial growth factor; βcatenin
INTRODUCTION
Colorectal cancer (CRC) is the third most common malignancy of the digestive tract and the fifth leading cause of cancer-related mortality in China[1,2]. The agestandardized incidence rate of CRC in China has increased from 12.8 per 100000 in 2003 to 14.2 per 100000 in 2017, while the mortality rate has risen from 5.9 to 7.4 per 100000. China has lower rates of CRC incidence and mortality than most developed countries, but has a higher case-fatality ratio (14.0%) and mortality/incidence ratio(52.1%)[3].
Treatment for CRC generally consists of surgery, adjuvant radiation, and chemotherapy as well as immunotherapy. Due to the low survival rate of CRC patients, there is an urgent need for new agents to combat this malignancy[4]. Plants are a rich source of various phytochemicals that may exert anti-oxidative, proapoptotic,anti-proliferative, anti-metastatic, and anti-angiogenic effects, depending on tumor type[5-8]. Sporamin is a Kunitz-type trypsin inhibitor that is found in the dicotyledonous plant, sweet potato (Ipomoea batatas), which belongs to the Convolvulaceae family. The tuberous roots contain 0.49%-2.24% crude sporamin protein on a fresh-weight basis[9,10]. Previous studies have identified sporamin as a potential anti-cancer agent against a number of malignant tumor cells, including HT29,HCT116, and SW480 colorectal cancer cells[11], TCA8113 tongue carcinoma cells[12]as well as PANC-1 and BxPC-3 pancreatic cancer cells[13].
The liver plays a vital role in the development of digestive tract cancers. It secretes a myriad of endocrine factors that may facilitate the growth and transformation of tumors, including β-catenin and vascular endothelial growth factor (VEGF). It is also the main metastatic target of CRC[14-16]. Thus, substances that can reduce the levels of tumor biomarkers in the liver may have the potential to become promising anti-cancer agents in the future. However, the effects of sporamin on the expression and secretion of tumor biomarkers in the liver are currently unknown. Therefore, in the present study, LoVo colorectal carcinoma cells were intraperitoneally xenografted into athymic BALB/c nude mice and sporamin was given orally to the mice to observe its effect on the growth of tumors, with a focus on changes in the structure and function of the liver, especially the expression and secretion of β-catenin and VEGF.
MATERIALS AND METHODS
Materials
Sporamin was extracted from sweet potatoes as previously reported[8]. Reagents used in these experiments were obtained from the following suppliers: SYBR Green and cDNA Reverse Transcription Kit, Thermo Scientific, Shanghai, China; TRIzol and diethyl pyrocarbonate, Invitrogen, Shanghai, China; anhydrous ethanol, chloroform,and isopropanol, Beijing Chemical Reagent Company, Beijing, China; Dulbecco's modified Eagle medium (DMEM), fetal bovine serum, and PBS buffer, Corning, NY,United States; penicillin-streptomycin mixture and trypsin, Keygen Biotech, Nanjing,China; VEGF and β-catenin ELISA kits, Mecenbio, Beijing, China; xylene, gradient ethanol, 1% hydrochloric acid ethanol, and chloral hydrate dry powder, Sinopharm Science and Technology, Beijing, China; distilled water, Experimental Platform of Capital Medical University, Beijing, China; neutral formalin fixative, Leagene, Beijing,China; 3% hydrogen peroxide, BSA, hematoxylin dyeing solution, neutral gum, DAB chromogenic reagent, and β-catenin polyclonal antibody, Solarbio, Shanghai, China;VEGF polyclonal antibody, Abcam, Beijing, China; HRP-conjugated goat anti-rabbit antibody, KPL, Wuhan, China; RNAlater RNA stabilization reagent, Sigma, St. Louis,MO, United States.
Cells
The colon cancer LoVo cell line was purchased from the Tumor Cell Bank of the Chinese Academy of Medical Sciences (Beijing, China). The cells were grown in DMEM high-glucose medium supplemented with 10% fetal bovine serum and 1%penicillin-streptomycin and maintained at 37 °C in a humidified incubator with 5%CO2. The medium was changed every 48 h. Single cell suspensions containing 5 × 106cells in 0.2 mL PBS were prepared during the logarithmic growth phase.
Animal experiments
All experiments were approved by the local Ethics Committee for Animal Research Studies at Capital Medical University, Beijing, China (animal experiment ethics review number: AEEI-2016-018).
Twenty-seven male BALB/c nude mice aged 4-6 wk old and weighing 13-15 g were purchased from Vital River Laboratory Animal Technology Co. Ltd. (Beijing, China)and maintained under specific pathogen-free conditions with free access to drinking water throughout the experiments. Animals were housed in a restricted access room under a 12-h/12-h light/dark cycle with a controlled temperature of 18-29 °C, daily range of temperature ≤ 3 °C, relative humidity of 40%-70%, airflow velocity ≤ 0.18 m/s, and room air pressure gradient of 20-50 Pa.
After one week of acclimation, animals were randomly divided into three groups,with nine animals in each group, and we verified that there was no significant differences in body weight between the groups. The groups were as follows: (1) Control group; (2) Vehicle group; and (3) Sporamin group. In the latter two groups, 5 × 106LoVo cells in 0.2 mL PBS were xenografted into the abdominal cavity of the mice.Mice in the control group were injected with an equal volume of PBS to mimic the operation in the other groups. Sporamin was dissolved in distilled water and given through intragastric infusion to the mice at a dose of 0.5 g/kg body weight/d. Mice in the control and vehicle groups were given the same amount of distilled water by intragastric infusion. The body weights of the mice were recorded every three days during the experiment. After three weeks of sporamin treatment, the mice were anesthetized with 10% chloral hydrate (0.1 mL/10 g body weight) and sacrificed by cervical dislocation. An autopsy of each mouse was carried out and the liver and the tumor nodes formed in the abdominal cavity were carefully counted, collected, and weighed. Then, the tissues samples were divided into three portions. Samples for pathological examinations were fixed in 10% neutral formalin and stored at room temperature. Samples for quantitative PCR (qPCR) were preserved in RNA stabilization reagent and stored at -20 °C. Samples for ELISA assays were directly stored at -80 °C in cryotubes.
Pathological examinations
Tissue samples fixed with formalin were subjected to paraffin embedding and sectioned at 5 μm for observation. Hematoxylin and eosin (HE) staining of the sections was performed to observe morphological changes in the tissues. After dewaxing and antigen retrieval, immunochemical assays were performed using specific polyclonal antibodies against VEGF and β-catenin. HRP-conjugated goat anti-rabbit antibody was then applied to the sections to label the primary antibodies. DAB chromogenic reagent and hematoxylin were then added and coverslips were added to sections after color development. The expression of VEGF and β-catenin in liver tissue was qualitatively analyzed with a high-magnification inverted fluorescence phasecontrast microscope (40×; DMIL, Leica, Germany) and photographs were taken.ImageJ was used for image analysis to compare the average optical density (AOD) of positive staining sites in each group.
Real-time fluorescent qPCR
Total RNA was extracted from the liver with TRIzol reagent. The mRNA was reversetranscribed into cDNA and SYBR Green PCR Master Mix was used to determine the transcriptional expression of specific genes. The primer sequences were as follows: βcatenin forward, 5’-TCT GAG GAC AAG CCA CAG GA-3’ and reverse, 5’-GCA CCA ATG TCC AGT CCA AG-3’; VEGF forward, 5’-CTT CAG CTC GCT CCT CCA CT-3’and reverse, 5’-CAG GCC TCT TCT TCC ACC AC-3’; β-actin forward, 5’-GTG CTA TGT TGC TCT AGA CTT CG-3’ and reverse, 5’-ATG CCA CAG GAT TCC ATA CC-3’. Amplification of the housekeeping gene beta-actin from the same samples was used as an internal control. Relative gene expression was calculated using the 2-ΔΔCtmethod[17].
ELISA
Liver samples were ground into homogenates and assayed according to the VEGF and β-catenin ELISA kit manufacturer’s instructions. The OD value of each sample well was measured with a microplate reader at a wavelength of 450 nm, and the concentrations of VEGF and β-catenin in the liver homogenates were calculated and presented in pg/mL. Reproducibility was evaluated in three independent experiments, and two standard curves were run on each plate.
Statistical analysis
Statistical analyses were performed using SPSS 21.0 software (IBM, Armonk, NY,United States). Image analysis was performed using ImageJ (https://imagej.nih.gov).Statistical charts were created using Prism5.0 (Graphpad, San Diego, CA, United States). All data are expressed as the mean ± standard deviation (SD). The differences between any two groups were tested by independent sample t-tests and comparisons of multiple groups were made by one-way ANOVA; Tukey’s test was used for multiple pairwise comparisons of means. A P-value < 0.05 was considered statistically significant.
RESULTS
Body weight, liver weig ht, and tumor burden
The body weight of the animals in all the three groups showed an upward trend as the experiment progressed. From day 9-15, body weight increased rapidly, but slowed from day 16-21. In comparison with the control and vehicle groups, it was noteworthy that the body weight of animals in the sporamin group was highest although there were no significant differences among these groups when the increase in body weight ceased on day 15 (P = 0.12; Figure 1). There were also no significant differences in liver weights or liver/body weight ratios among the three groups (Table 1).
All of the nude mice that were inoculated with cancer cells developed tumor nodules in their abdominal cavities. Figure 2 shows that the tumors grew mainly on the mesentery instead of the intestine. They were rough, hard, and grayish white nodules differing in number and volume. Compared to the vehicle group, the total weight of the tumor nodules in the sporamin group was reduced by 51.15% (95%confidence interval: 0.16-0.31) and the total number of tumors was reduced from 93 to 55 (Figure 3). Compared with the vehicle group (0.44 ± 0.10 g), the mean tumor weight in the sporamin group (0.26 ± 0.15 g) was significantly reduced (P = 0.04).
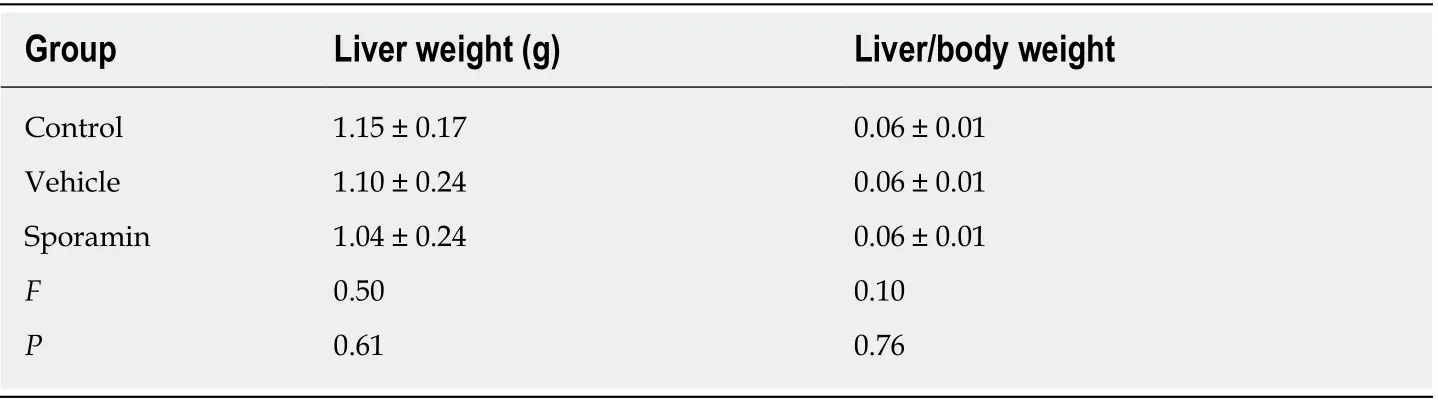
Table 1 Liver weights and the liver/body weight ratios of the BALB/c nude mice
Pathological examinations
HE staining of liver histological sections showed that, compared to the control group,the size of hepatocytes in the vehicle group was enlarged, and nuclei were also enlarged and stained deeply blue. The cytoplasm displayed a basophilic change. Cell membranes and the borders between them became blurred. As a result, the hepatic sinusoid was compressed by the enlarged hepatocytes and the structure of the hepatic plate became irregular. There were also many lymphocytes which had infiltrated into the hepatic tissues, but no metastatic tumor cells were found. Compared to the vehicle group, the histomorphology of livers from animals in the sporamin group was similar to that of the control group with a relatively small cell size, clearer hepatic plate, and a reduced degree of blue staining of the cytoplasm, indicating that sporamin had restored the normal structure of the liver in these animals (Figure 4).
Immunohistochemistry
To investigate functional changes in the liver, antibodies against β-catenin and VEGF were applied to the liver sections to show the levels and intracellular locations of these tumor biomarkers. After antibody incubation, the nuclei of the liver cells were stained blue and biomarkers were stained brownish yellow or light yellow. As shown in Figure 5, high concentrations of β-catenin were found in the cytoplasm and also translocated to the nucleus, indicating that it had bound to its target genes and had initiated the transcription process. Quantitative analysis of the AOD of the positively stained areas in the liver tissue sections showed that, compared with the vehicle group (47.29 ± 9.10), the expression of β-catenin was significantly reduced by sporamin to 26.14 ± 1.72 (P = 0.003; Figure 6). In line with the increase in β-catenin expression and nuclear translocation in the vehicle group, the expression of the angiogenic factor VEGF, which is also a downstream target of β-catenin, was increased in the cytoplasm of hepatocytes in the vehicle group. Similarly, expression of the VEGF protein was also reduced after sporamin intervention from 20.78 ± 2.06 in the vehicle group to 15.80 ± 1.09 in the sporamin group (P = 0.021; Figure 6), sug-gesting that sporamin had an anti-angiogenic effect in these animals.
ELISA
The concentrations of β-catenin and VEGF protein in liver tissue were assessed by ELISA. Figure 7 shows that the concentration of β-catenin increased, albeit nonsignificantly, from 134.42 ± 22.04 pg/mL in the control group to 143.33 ± 5.06 pg/mL in the vehicle group (P = 0.35). However, the concentration of VEGF in liver tissue was significantly increased from 132.05 ± 7.96 pg/mL in the control group to 158.73 ±6.23 pg/mL in the vehicle group (P = 0.00007). Compared with the vehicle group, the concentration of β-catenin after sporamin intervention decreased to 109.07 ± 9.65 pg/mL (P = 0.00002). The concentration of VEGF also decreased to 150.90 ± 10.38 pg/mL but it was not significant (P = 0.14). These results are consistent with changes observed in the immunohistochemistry analysis.
qPCR
To further verify the effects of sporamin on the expression of β-catenin and VEGF at the transcriptional level, qPCR was conducted to detect the relative abundance of these mRNAs in the liver samples. Figure 8 shows that compared with the control group, relative expression of β-catenin mRNA in the vehicle group was significantly increased (P = 0.0004), but the increase was not significant for VEGF (P = 0.26).Compared with the vehicle group, the relative abundance of β-catenin and VEGF mRNA in the sporamin group was significantly reduced to 71% ± 1% (P = 0.000001)and 23% ± 7% (P = 0.00002) of the vehicle group levels, respectively.
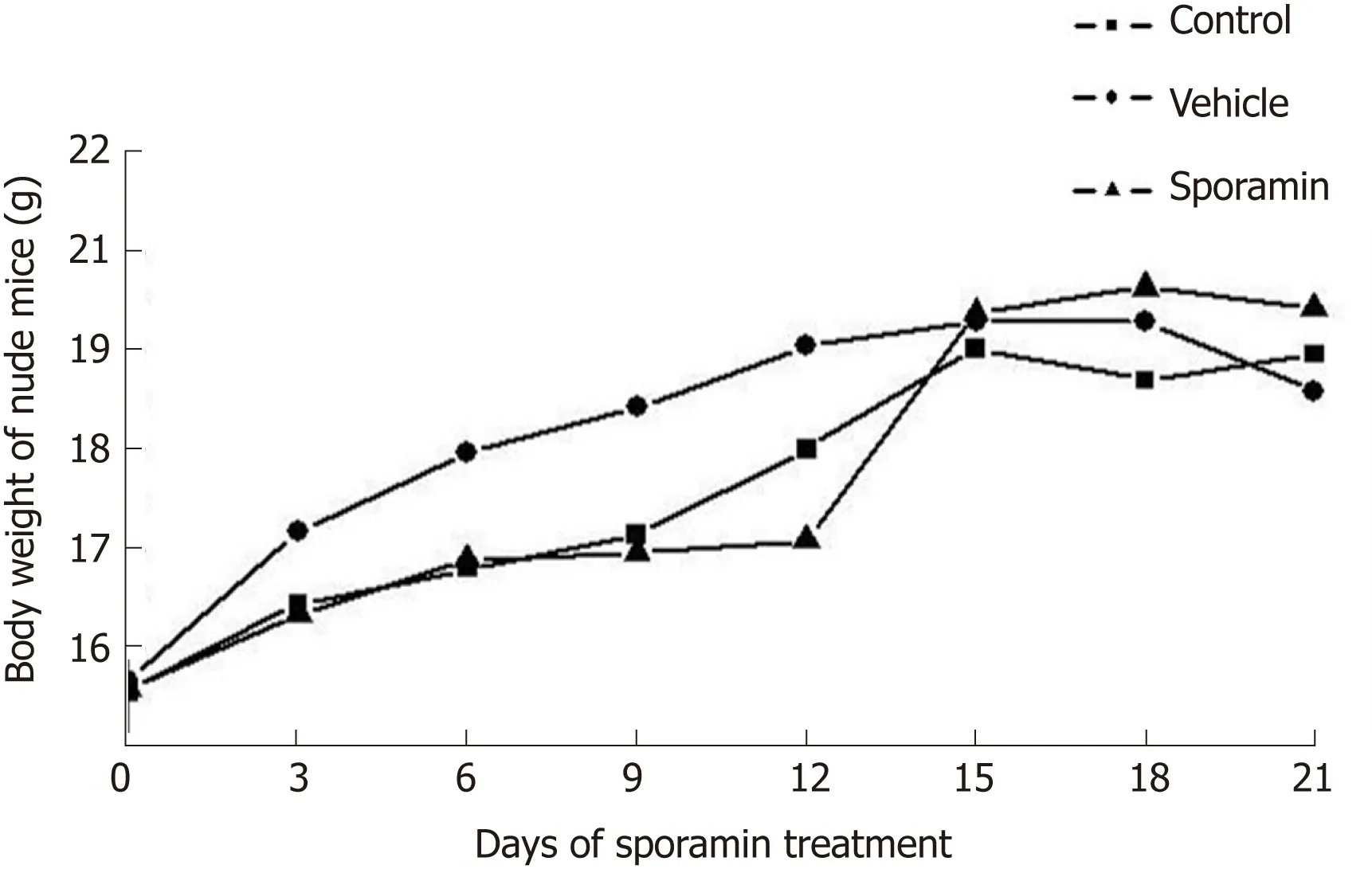
Figure 1 Body weight changes during sporamin treatment.
DISCUSSION
CRC is one of the most common malignancies of the digestive tract in both developing and developed countries[18]. Metastasis and recurrence of the primary tumor are the main reasons for the high mortality rate of this disease. At present, chemotherapy still has severe side effects, which limits its use in many circumstances. In recent years, many studies have indicated that phytochemicals may be a promising source of new anti-cancer agents against CRC which will play an important role in chemoprevention of the disease (for review see[19]). In the current study, compared with the vehicle group, the number and total weight of tumor nodules formed in the abdominal cavities of the mice were significantly reduced by treatment with sporamin, a Kunitz type trypsin inhibitor obtained from the sweet potato. This is in line with previous studies which showed that sporamin can suppress the growth of a variety of cancer cell lines including human esophageal squamous cell carcinoma cells[20], human pancreatic cancer cells[13], and human tongue carcinoma cells[12], both in vitro and in vivo.
Considering that the liver is usually the first target organ of CRC metastasis, the present study mainly focused on changes in the structure and function of the liver in the tumor-bearing mice. The liver is the largest endocrine gland in the body which can secrete a great number of hormones, growth factors, and cytokines and plays a crucial role in the development and transformation of many malignant tumors. The liver is also the most common anatomical site for hematogenous metastases of CRC, which are one of the most difficult and challenging obstacles in the treatment of CRC[21]. In our study, although the weights of the body and the liver and the ratio of liver weight to body weight were not significantly different among the control, vehicle, and sporamin groups, we found that sporamin increased body weight and reduced liver weight compared with vehicle treatment, implying that sporamin was possibly beneficial for the general health status of the mice. This effect may be partially attributed to the nutritional effects of sporamin because it is a dietary protein with many biological activities[22].
In accordance with previous findings that colon cancer cells can induce the liver to synthesize and secrete a variety of hormones, growth factors, and cytokines, which facilitate the growth of the tumor and induce a systematic inflammatory status[23], our studies showed that after intraperitoneal tumor cell inoculation, the histomorphology and function of the liver were all significantly changed . In contrast, it was noteworthy that, compared with the vehicle group, sporamin demonstrated the ability to restore the normal structure and function of the liver. Hepatocyte enlargement,nuclear pyknosis, cell membrane blurring, and blue staining of the cytoplasm were all improved, suggesting that sporamin had attenuated protein synthesis within the cell and alleviated the hypertrophic status of the hepatocytes. Functionally, the accumulation of β-catenin in the cytoplasm as well as its translocation to the nucleus of the hepatocytes was both significantly attenuated by sporamin. As is well known, the activation of the Wnt/β-catenin signaling pathway plays a key role in the development of CRC[24]. As a transcriptional activator, the translocation of β-catenin from the cytoplasm to the nucleus initiates the transcription of a number of target genes[25]which participate in the malignant transformation of CRC[26]. Therefore, any agents that can reduce the level of β-catenin within the tumor cells and hepatocytes may exhibit a potent effect against CRC. For example, in ade-nomatous polyposis coli(APC) tumor suppressor gene-mutated APCmin/+mice, the formation of colorectal adenomas is inhibited by suppressing the activation of the Wnt/β-catenin signaling pathway[27]. Therefore, our findings suggest that sporamin may partially exert its effect by inhibiting the synthesis and function of β-catenin in the liver.
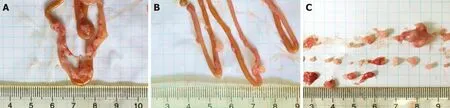
Figure 2 Establishment of a colorectal cancer in vivo model. Photos taken of intestines from the vehicle group (A) and sporamin-treated group (B). C: Tumor shapes are shown.
VEGF is one of the most potent angiogenic factors that can be synthesized by tumor cells as well as hepatocytes after stimulation by various environmental factors such as hypoxia. VEGF is also a downstream target of the β-catenin pathway, and expression of VEGF is usually positively correlated with the expression of β-catenin in tumor cells and with the progression of the tumor[28]. Consequently, if the expression of βcatenin in tumor cells is reduced, the expression of VEGF usually decreases as well[29].Our results are consistent with these previous findings and show that sporamin has an anti-angiogenic effect against CRC. As to the specific mechanisms by which sporamin inhibits the expression of β-catenin and VEGF as well as signaling events during this process, further investigation will be required.
In conclusion, our study suggests that sporamin can suppress the growth of xenografted colorectal tumor nodules in mice by restoring the normal structure of the liver and downregulating the ex-pression and secretion of β-catenin and VEGF in the liver. These anti-cancer effects of sporamin against CRC are closely associated with its inhibitory effect on these tumor biomarkers. Further studies are warranted to elucidate the corresponding signal transduction events mediating this process.
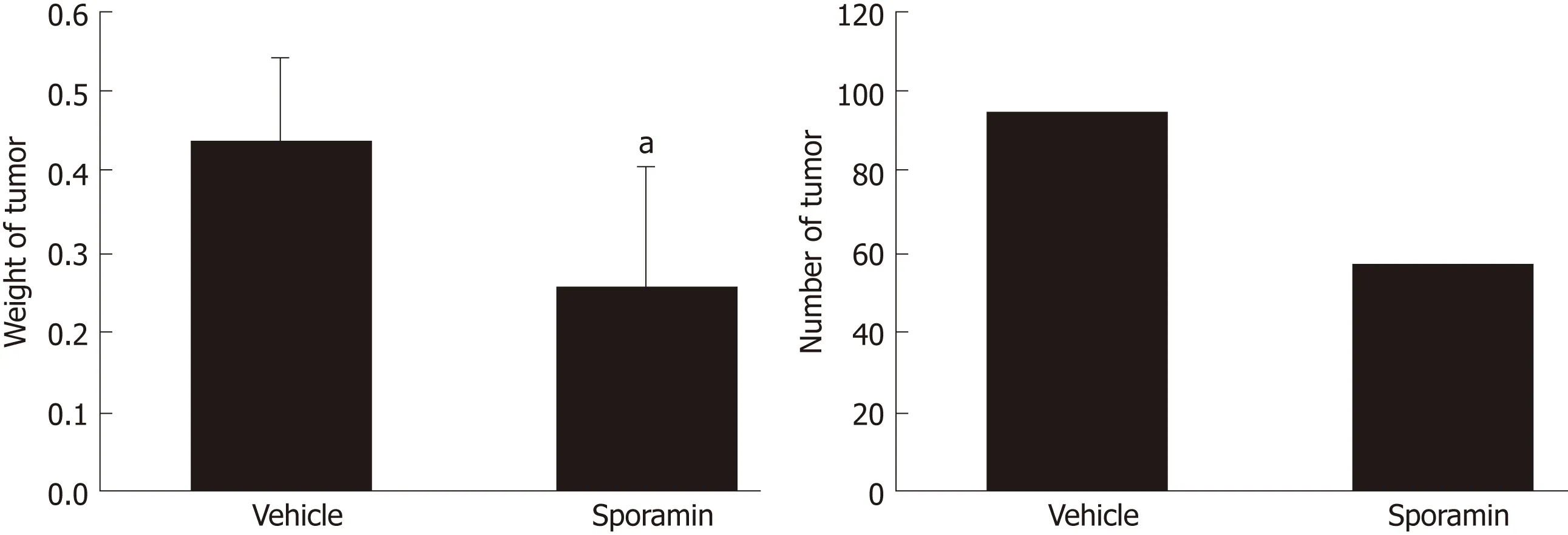
Figure 3 Changes in the weight and number of tumors following sporamin administration.aP < 0.05 vs vehicle.
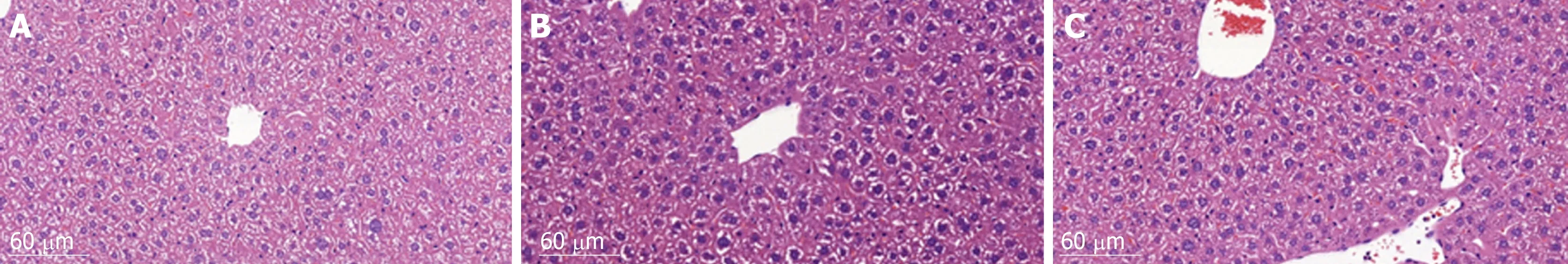
Figure 4 Liver histological changes after sporamin treatment. Photomicrographs of hematoxylin and eosin staining of liver sections from the normal control group(A), vehicle group (B), and sporamin group (C). Magnification, 20×.
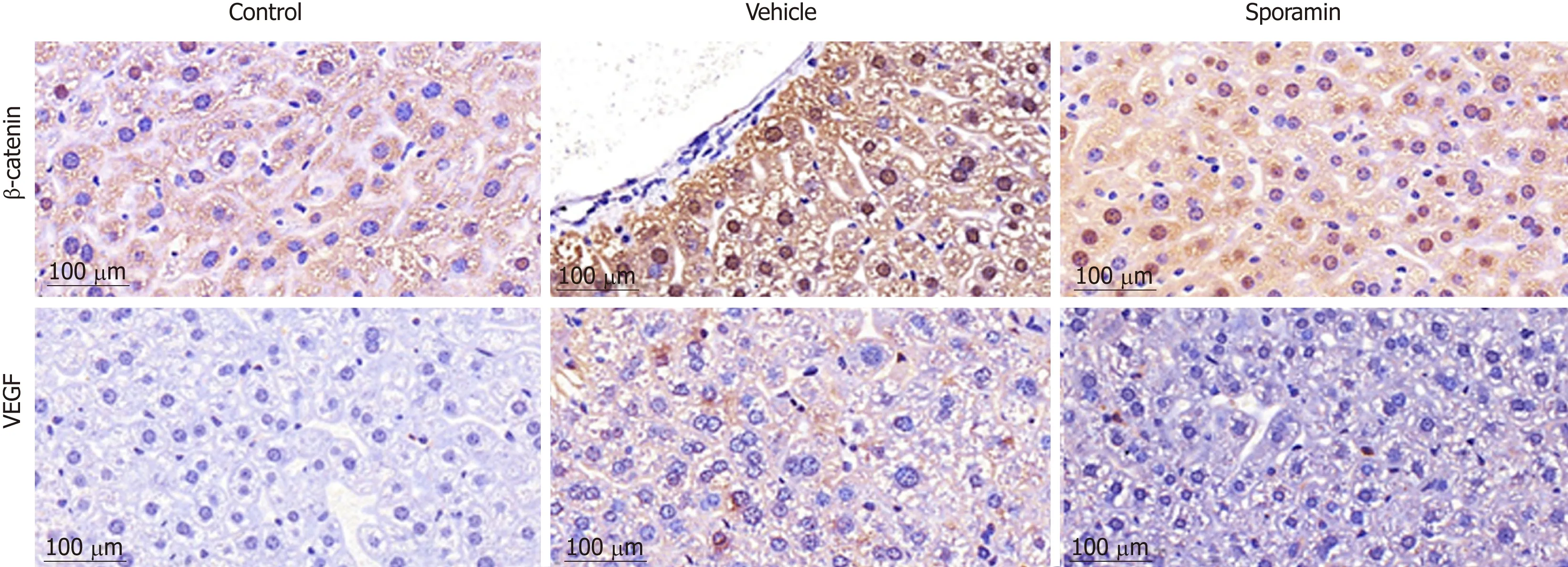
Figure 5 Immunohistochemical staining for vascular endothelial growth factor and β-catenin in the liver of nude mice after sporamin treatment.Photomicrographs of liver sections from the control group (A), vehicle group (B), and sporamin group (C). Magnification, 40×. VEGF: Vascular endothelial growth factor.
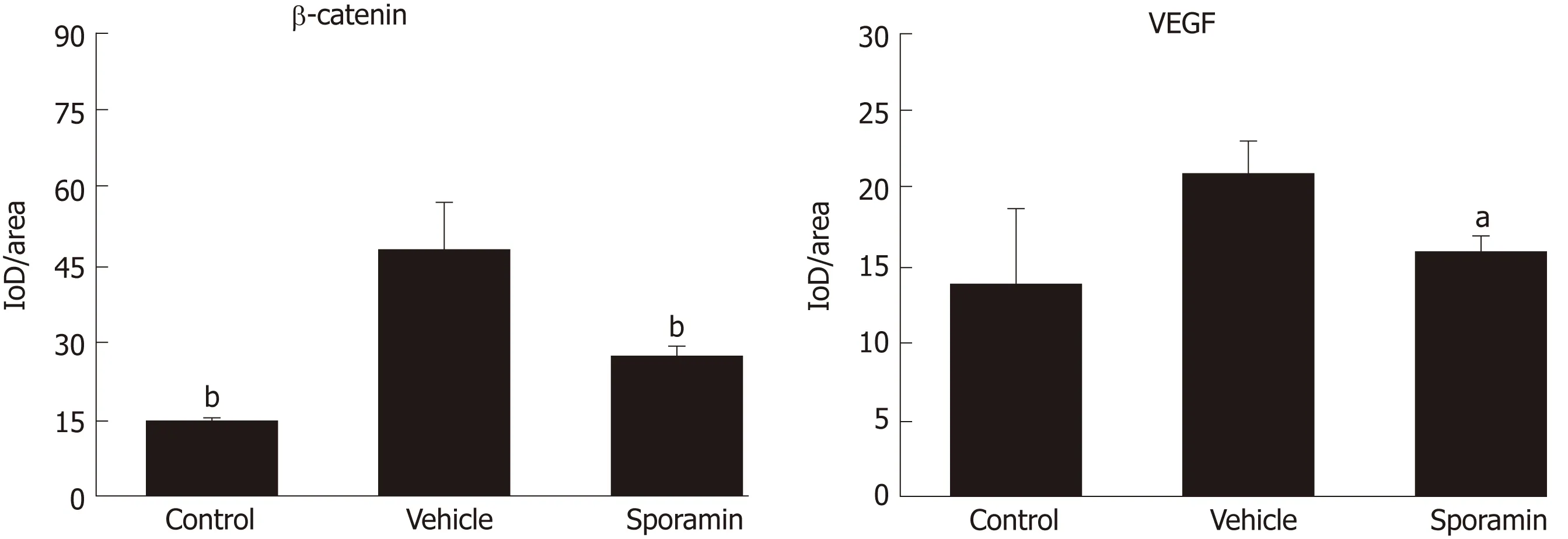
Figure 6 Average optical density of vascular endothelial growth factor and β-catenin in the liver of nude mice after sporamin treatment. Measurements were made using ImageJ; n = 3 per group. aP < 0.05 vs vehicle, bP < 0.01 vs vehicle. VEGF: Vascular endothelial growth factor.
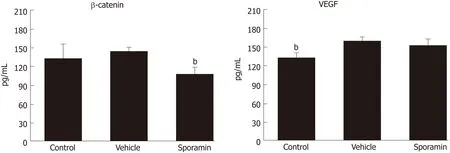
Figure 7 Concentrations of β-catenin and vascular endothelial growth factor in the liver of nude mice after sporamin treatment.bP < 0.01 vs vehicle. VEGF:Vascular endothelial growth factor.
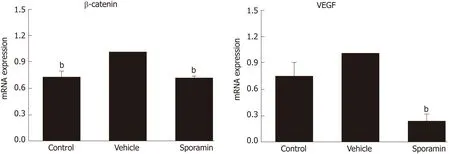
Figure 8 Relative mRNA expression of β-catenin and vascular endothelial growth factor.bP < 0.01 vs vehicle.
ARTICLE HIGHLIGHTS
Research background
Colorectal cancer (CRC) is the third most common malignancy of the digestive tract and the fifth leading cause of cancer-related mortality in China. Sporamin, a Kunitz-type trypsin inhibitor isolated from sweet potato, is a potential anti-cancer agent with activity against a number of malignant tumor cells in vitro. The liver secretes a myriad of endocrine factors that may facilitate the growth and transformation of tumors in the development of CRC.
Research motivation
Sporamin as a potential anti-cancer agent against a number of malignant tumor cells, including HT29, HCT116, and SW480 colorectal cancer cells, TCA8113 tongue carcinoma cells as well as PANC-1 and BxPC-3 pancreatic cancer cells. However, the effects of sporamin on the expression and secretion of tumor biomarkers in the liver are currently unknown. Therefore, in the present study, LoVo colorectal carcinoma cells were intraperitoneally xenografted into athymic BALB/c nude mice and sporamin was given orally to the mice to observe its effect on the growth of tumors, with a focus on changes in the structure and function of the liver, especially the expression and secretion of β-catenin and vascular endothelial growth factor (VEGF).
Research objectives
To investigate the effects of sporamin on liver morphology and biomarkers of xenografted CRC in the liver of BALB/c athymic mice.
Research methods
Twenty-seven male BALB/c nude mice were randomly divided into control, vehicle, and sporamin groups. Mice in the latter two groups were intraperitoneally xenografted with LoVo colorectal carcinoma cells and intragastrically infused with saline or sporamin (0.5 g/kg body weight/d), respectively, for 3 weeks. Hematoxylin and eosin (HE) staining of the sections was performed to observe morphological changes in hepatic tissue and real-time fluorescent quantitative PCR and enzyme-linked immunosorbent assays (ELISA) were used to measure the expression of β-catenin and VEGF in the liver.
Research results
Sporamin significantly reduced the number and weight of tumor nodules formed in the abdominal cavity. Compared with the vehicle group, the mean tumor weight (± SD) in the sporamin group was significantly reduced (0.26 ± 0.15 g vs 0.44 ± 0.10 g) and the total number of tumors decreased from 93 to 55. HE staining showed that enlargement of the nucleus and synthesis of proteins within hepatocytes, as well as infiltration of inflammatory cells into the liver, were attenuated by sporamin. Immunohistochemical staining and ELISA showed that the concentrations of β-catenin and VEGF in the liver were significantly reduced by sporamin.Compared with the vehicle group, the expression of β-catenin measured in integrated optical density units per area was reduced in the sporamin group (47.29 ± 9.10 vs 26.14 ± 1.72; P = 0.003).Expression of VEGF was also reduced after sporamin intervention from 20.78 ± 2.06 in the vehicle group to 15.80 ± 1.09 in the sporamin group (P = 0.021). The secretion of VEGF and βcatenin in the liver was also assessed by ELISA, which showed that the concentration of VEGF in liver tissue increased significantly from 132.05 ± 7.96 pg/mL in the control group to 158.73 ± 6.23 pg/mL in the sporamin-treated group (P = 0.00007). Compared with the vehicle group, the concentration of β-catenin decreased from 134.42 ± 22.04 pg/mL to 109.07 ± 9.65 pg/mL after sporamin intervention (P = 0.00002). Quantitative PCR (qPCR) indicated that compared to the vehicle group, relative mRNA expression of β-catenin and VEGF in the liver of the sporamintreated group was significantly reduced to 71% ± 1% (P = 0.000001) and 23% ± 7% (P = 0.00002),respectively, of the vehicle group levels.
Research conclusions
Sporamin down-regulates the expression and secretion of β-catenin and VEGF in the liver, which subsequently inhibits the transcription of downstream genes involved in cancer progression and angiogenesis.
Research perspectives
Our study suggests that sporamin can suppress the growth of xenografted colorectal tumor nodules in mice by restoring the normal structure of the liver and downregulating the expression and secretion of β-catenin and VEGF in the liver. These anti-cancer effects of sporamin against CRC are closely associated with its inhibitory effect on these tumor biomarkers. Further studies are warranted to elucidate the corresponding signal transduction events mediating this process.
杂志排行

World Journal of Gastroenterology的其它文章
- Advanced imaging in surveillance of Barrett’s esophagus: Is the juice worth the squeeze?
- Fate plasticity in the intestine: The devil is in the detail
- Revisiting the liver’s role in transplant alloimmunity
- Hepatocellular carcinoma: Therapeutic advances in signaling,epigenetic and immune targets
- Hepatocellular carcinoma: Mechanisms of progression and immunotherapy
- Epidemiology of hepatitis E in South-East Europe in the "One Health" concept