植物光信号转导
2019-07-03李秀坤许冬清
李秀坤,许冬清
南京农业大学 农学院 作物遗传与种质创新国家重点实验室,南京 210095
太阳光是植物能量的主要来源。除此之外,光还作为一个重要的环境信号因子影响和调控植物的诸多生长发育和生理过程。自20世纪90年代以来,植物光生物学家借助分子生物学和分子遗传学的发展和各种新的技术方法,对植物光信号转导进行研究并有突破性的进展。植物接受光信号并传递至下游,进而作出适时的应答反应,以适应时刻变化的光环境。植物是固生生物,从种子落地发芽开始,就会在一个固定的地点完成整个生命周期。但是,植物并不像我们看到的那样静止不动,在微观层面上可谓是瞬息万变。不同光环境下,植物启动精密的光信号转导系统以应对不同的光质、光强、光照时间和方向,确保其自身的健康生长。
1 植物识别光信号的物质——光受体
阳光普照一切生物,为大自然带来生机。所谓 大海航行靠舵手,万物生长靠太阳 。自1906年美国科学家Garner和Allard发现光对植物生长发育的影响开始,全世界的科学家就从未停止过光对植物作用的研究。
在数万年的进化过程中,植物为了感知和识别光信号已经进化出几类不同的光受体。它们是识别280~315 nm 的 UV-B 信号的紫外光受体 UVR8[1-2]、吸收波段为315~500 nm 的 UV-A和蓝光受体隐花色素 1 和 2(CRY1和CRY2)以及向光素 1 和 2(Phot1和Phot2)[3-5],而光敏色素(PHYA~PHYE)主要吸收600~750 nm的红光和远红光[6](图1)。其中PHYA是吸收远红光信号的主要光受体,PHYB是识别红光信号的主要光受体。光敏色素在吸收波长为670 nm的红光后可以转化成有活性的光受体Pfr,而在吸收730 nm的远红光后转换成无生物学活性的Pr状态[7]。这些光受体的发现,对于植物光生物学的发展具有里程碑意义。全世界多家致力于光信号转导研究的实验室,协同参与了这些重要的研究项目。斯坦福大学的Winslow Briggs院士发现向光素,并解释了植物的向光性现象[3]。加州大学伯克利分校的Peter Quail院士克隆了第一个光敏色素[6]。宾夕法尼亚大学的 Anthony R. Cashmore院士和加州大学洛杉矶分校的林辰涛教授领导的研究团队分别克隆了隐花色素CRY1和CRY2[4-5]。UV-B受体由瑞士日内瓦大学Roman Ulm教授实验室首先发现[1]。有人会问:为什么没有识别500~600 nm波段的光受体?答案是:这一波段为绿光,在植物体内是没有绿光受体的。绿光可以在红光受体和蓝光受体的介导下,参与植物生长发育的过程,如向光性反应、茎的伸长、叶的生长、气孔的开闭和其他器官的生长等。人类肉眼只能看到可见光部分,一般为400~760 nm之间。由此看来,植物比人类感受光的能力要更强。这些光受体在识别不同波长的光信号后,迅速地在植物细胞内启动不同的生物学事件,将信号传递至下游,并通过影响大量基因的表达而调控不同的生理和生长发育过程。
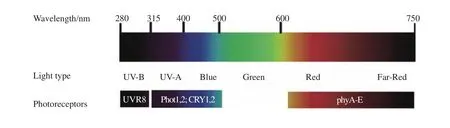
图1 模式植物拟南芥中的光受体
2 光信号转导的重要核心——COP/DET/FUS
植物通过这些光受体精确分辨太阳光不同的波长、方向、强度和周期等信息变化,经由复杂的信号感受、传递和响应,完成信号转导过程。一方面植物利用光能进行光合作用,将光能转化成有机能;另一方面,通过一系列光调节基因来调控种子的萌发、幼苗的光形态建成、避荫性、叶绿体和小叶的运动、开花的诱导以及很多细胞核内和叶绿体内基因的表达等。为了揭示其中的奥秘,植物光生物学家做了很多有意思的研究,以期揭示这个光信号转导通路和植物响应光的分子机理。最终科学家们通过遗传筛选和表型鉴定获得了cop/det/fus这样一组对光信号具有多效性的突变体[8](图2)。这些突变体在黑暗下都表现出组成型光形态建成的表型,即这些基因单独的功能缺失均可以导致植物幼苗在黑暗下就模拟光下生长的过程。COP1是COP/DET/FUS系统中一个典型的代表,是光形态建成的核心抑制因子。它实际上是将光形态建成的促进因子进行降解而行使抑制光形态建成的机制,通过多泛素化光形态建成核心促进因子HY5、HYH、PHYA、BBX21、BBX22、HFR1、PIL1、PAR1和PAR2等,而促进它们通过26S蛋白水解酶体降解[8-9]。在光照条件下,一方面有活性的光敏色素受体从细胞质进入细胞核后会与 COP1 或者 SPAs 等蛋白互作来解离 COP1-SPAs 复合体,进而抑制COP1的E3泛素连接酶活性,而它的活性降低就会使得植物表现出光形态建成的表型;另一方面大部分 COP1 以目前还不太清楚的机制出核,最终使得细胞核内光形态促进因子迅速积累,进而促使幼苗进行光形态建成[10-11]。DET是Deetiolation的简称,也就是去黄化。植物在黑暗下生长,会出现黄化苗,见光以后则会合成叶绿素转为绿色。 FUS 则是Fusca的简称。Fusca是拉丁语,深紫色的意思,主要是因为这些突变体的种子中花青素大量积累,而呈现出深紫色。其中cop系列的突变体和det系列的突变体最后都被定位是相应的fusca系列突变体的等位基因,如cop1与fusca1是等位的,det1与fusca2是等位的。COP/ DET/ FUS 位于光受体的下游,处在光信号通路的中心,它们主要通过调节下游的光形态建成促进因子的蛋白水平来调节幼苗的光形态建成。
COP1是其中一直较为受关注的成员。作为E3 泛素化连接酶,其通过降解参与植物光形态建成的促进因子以及调节开花和生物钟节律的一系列调控因子等,来参与植物生长发育的诸多过程。1991年邓兴旺(后来当选为美国科学院院士)首先鉴定了拟南芥cop1突变体[12]。次年,他成功地克隆了拟南芥COP1基因,这一研究成果发表在《细胞》(Cell)杂志上[13]。随后,邓兴旺教授领导的研究团队通过系统性的遗传和生物化学研究鉴定和证明了CP/DET/FUS 所组成的3个E3泛素化连接酶复合体位于光受体下游,并处于光信号转导途径的核心位置。这三大复合体在黑暗下位于细胞核内并具有高度的生物学活性,降解了大量的蛋白底物,从而确保植物下胚轴或茎迅速伸长,以及双子叶植物子叶闭合并形成弯钩;而见光以后,这些复合体的活性被多种不同分子调控机制所抑制,使得下游底物在细胞核内得以积累并促进幼苗生长[8-9]。这些有趣的发现,不仅完善了人们对植物光信号转导的理解,也推动了针对这一调控系统在动物和人类中的研究。这是因为COP/DET/FUS系统在动物和人类中也具有E3泛素化连接酶活性,参与和调控肿瘤发生等诸多生理和病理的发生和调控过程。

图2 野生型和cop1突变体在黑暗下和光下生长的表型
3 光信号转导中的转录因子
光信号影响植物体内将近3 000多个基因的表达,而直接控制这些基因表达的主要是一类受光调控的转录因子,其中HY5是这一途径最为关键和最具有代表性的一个转录因子。HY5可以直接或者间接地结合在植物体内约1/3基因的启动子区域而调控它们的表达[14]。这些基因的改变直接影响着植物受光调控的生长发育过程,其中就包括光形态建成。在黑暗下,HY5是被COP1所泛素化而降解的;而在光下,积累的HY5就具有调控下游目的基因表达的能力[15]。在转录和蛋白水平,HY5受到光受体和COP/DET/FUS 的调控,证明HY5位于这一调控体系的下游,也就是说光信号-光受体-COP/DET/FUS系统通过调控HY5而影响植物体内大量基因的表达,从而最终调控植物的光形态建成。hy5突变体最初也是在邓兴旺教授的实验室通过遗传筛选获得[16]。随后,植物光生物学家借助于分子生物学和生物信息学的技术手段,阐明HY5在植物光信号转导途径中详细的工作机制,并确立它在这一转导途径中的重要位置。
4 光信号转导的微视角
拟南芥幼苗利用光受体感受光信号后,主要通过影响蛋白与蛋白的互作、蛋白的核质定位和蛋白的稳定性等生物学事件将信号传递下去。在光信号通路中,光影响蛋白与蛋白互作比较常见,如光敏色素被激活入核后可与 PIFs 和SPAs 等蛋白互作,受蓝光激活的 CRY2 与 CIB1和 SPAs 可以相互作用,以及紫外光照后二聚的UVR8 解离成 UVR8 单体后能够与 COP1/SPAs复合体相互作用等[10]。
在黑暗条件下,没有活性的光敏色素受体位于细胞质内,而细胞核里的 PIFs蛋白则以同源或者异源二聚体的形式结合在光调控基因的启动子上,并调控这些基因的表达,进而抑制植物的光形态建成。另外,位于细胞核内的COP1 能够降解光形态建成的促进因子如 HY5、 LAF1、HFR1、PAP1和PAP2等。HY5是光形态建成的正向调节子,HFR1参与避荫反应,PAP1和PAP2是花青素生物合成的正调控因子,而CO是光周期介导的促进开花的关键因子。这些关键因子控制了植物体内大约1/3基因的表达[10]。光受体PHYA、PHYB和CRY2也被COP1/SPA蛋白复合体降解,抑制光形态建成,表明COP1反馈调控光受体。在光照条件下,光敏色素迁移至细胞核里,能够与PIFs相互作用,并以一种未知的方式磷酸化PIFs而被降解[17];同时,光敏色素从细胞质进入细胞核后会与 COP1 或者 SPAs 等蛋白形成异聚体来解离 COP1-SPAs 复合体,进而抑制COP1-SPAs活性[10-11]。在长时间的光照下,大部分 COP1 以目前还不太清楚的机制迁移出细胞核,最终使得细胞核内光形态促进因子迅速积累,进而促使植物体内1/3基因的正常表达而促进光形态建成。因此,光受体-E3泛素化连接酶复合体-转录因子调控体系是植物光信号转导系统中的核心调控开关。
5 结束语
不同类别的光受体和COP/DET/FUS的发现和鉴定极大地加深了人们对光信号转导系统的认识,并且绘制了一个类似线性的光信号转导途径。COP/DET/FUS组成了3个不同的E3泛素连接酶复合体,降解诸多的下游底物。这些调控因子在包括人类在内的其他真核生物中也是保守的,并且具有类似的生物化学活性。除了调控光形态发生外,COP/DET/FUS蛋白在许多其他生物学过程中也起着关键作用,其在植物或者哺乳动物中的功能完全丧失,均导致致死的表型。基于黑暗下到光下植物幼苗形态建成的分子机理研究,已经揭示了一个较为清晰的光信号转导途径,但是光信号转导系统如何在调控植物其他的生长发育过程(例如气孔发育、成株形成等)中行使作用,以及与其他环境因子和不同植物激素信号途径的整合调控,将是植物光生物学家需要持续付诸努力的研究领域和方向。
