夜间人工照明对生物节律的影响
2019-06-25黄大明朱文迪刘光KevinJ.Gaston
黄大明 朱文迪 刘光 KevinJ.Gaston

摘 要:夜晚使用人工照明,在给人类带来很多好处的同时,也成为了改变很多生物活动节律的诱因,破坏了生物赖以生存的自然日光周期、季节光周期和月光周期。本文回顾了夜间人工照明影响生物节律的证据。虽然案例比较分散,但是考虑到物种和环境的多样性,这些影响很有广度。事实上,推测绝大多数人工夜间照明对生物的影响,是对生物节律的影响。这也支持了夜间人工照明对生态系统的普遍显著性影响,因此应该重视全球范围内直接照明和人工白昼的迅速扩张,而且需要采取广泛的措施缓解危害。
关键词:昼夜节律;照度;光周期;人工白昼;光谱
中图分类号:Q958 文献标识码:A 文章编号:1671-2064(2019)09-0184-07
0 引言
生物学中,节律无处不在。无论是基因表达、新陈代谢、生理活动或者机体运动、摄取营养,繁殖,正常的节律往往对于生物适应和生存起着非常重要作用。自然光周期,通常是生物感知到的最強烈和最可预测的环境波动[47]。因此光周期也是影响生物节律最重要的环境诱因,包括受昼夜节律调节的日常活动(如酶活、气孔开闭和气体交换、光合作用、花朵开闭、花香外溢、鸟鸣、觅食、运动、睡眠、暗修复和恢复),也包括依赖昼长或光周期变化的季节性活动(如发芽、爆芽、开花、生长、滞育、冬眠、蜕皮、繁殖、迁徙、衰老)。事实上,日周期、年周期和月光周期的多种组合主导并调节了大多数有机体的生命活动。
人们多次发现人为压力对生物多样性的影响,是通过影响生物生活史中的特定阶段发生的时间进行的。例如,气候变化正在导致食物供应与作物繁殖周期错配,城市化导致了生物种群呈现超前的物候现象[52]。在许多情况下,自然节律的人为影响,源于生物用于节律判断的环境诱因发生了变化。因此,人工照明对自然光周期的直接影响,对理解人类活动的生态影响至关重要。
夜间人工照明正日益侵蚀并改变自然光周期。人工照明有多种来源,包括路灯、建筑和车辆。从短期脉冲式照明(如过往车辆的前灯)到持续性夜间照明(如路灯),从窄带发射光谱到宽带发射光谱,从低发光强度到高发光强度,以及从局部的集中照明到遍布的人工白昼,人工照明呈现出各种各样的形式。
夜间人工照明的生物影响已有多年的文献记录,实验已明确了它对生物健康和幸福感的深刻影响[45]。近十年来实地研究也发现了很多案例,证明夜间人工照明对个体生理和行为、物种丰盛度和分布、生态相互作用、群落构成以及生态系统功能有影响。这些文章记录的影响有的是通过现场直接测量,有的是在此前未受人工照明波及的地区引入路灯进行实验,也有的是模拟路灯发光进行实验[9,26,45,54,57,92,98]。研究发现这些影响广泛存在于多种物种中,包括微生物、植物、软体动物、昆虫、鱼类、两栖动物、爬行动物、鸟类和哺乳动物[45]。然而,究竟夜间人工照明的生物影响,多大程度上取决于生物节律的改变,还不清楚。
本文探讨夜间人工照明对野生生物节律的影响。而不是具体的分子、细胞或者生理机制[5,80,100]。文中采用勒克司为单位描述光照强度。这是一种基于人的明视觉的单位面积光通量的单位,因此不能反映不同光谱对生物影响的相对效果。但是由于其广泛用于环境测量、人工照明系统的研发和改良,使用勒克斯能够确保与照度的直接关联。
1 自然光周期
影响生物节律的自然光周期主要有三种形式:日光周期、季节光周期和月光周期。
1.1 日光周期
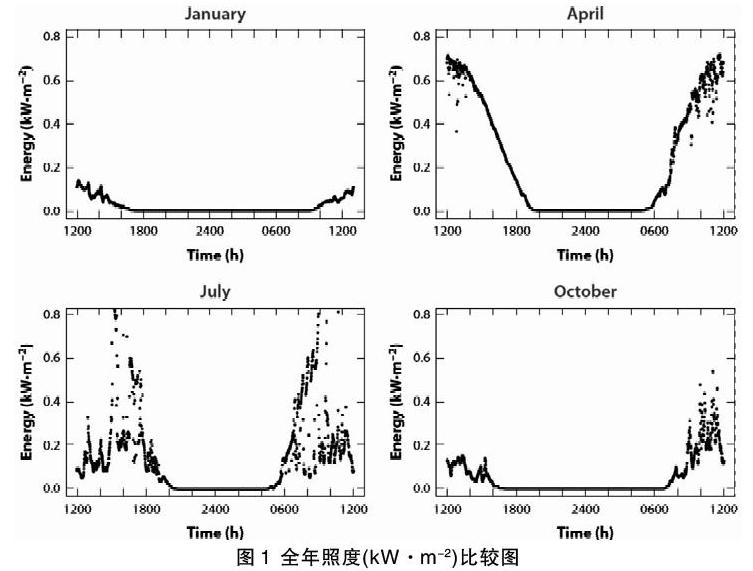
地球绕其轴线的自转将时间划分成规律的昼夜周期。在白天,来自太阳的入射光强度可达约100,000lx,而晴朗的夜晚星光约为0.001lx、月光约为0.1lx(满月),总跨度(以勒克司为单位)约六到八个数量级[45]。昼长是日出和日落之间的时间,日出与日落时太阳的中心均在地平线以下,昼长是这段过程的持续时间。白天的光照强度跨越了三个数量级[60]。虽然还有其他定义,一般认为暮曙光是日落或日出中太阳的几何中心在地平线以下6°的时段(民用暮曙光),也是太阳的几何中心在地平线以下6°和12°之间的时段(航海暮曙光),或是太阳的几何中心在地平线以下12°和18°之间的时段(天文暮曙光)。夜晚是暮光和曙光之间,太阳不可见的那段时间。在暮曙光时光照强度的变化速率最大,在黄昏时随着太阳高度角的减小而减少,在黎明时随着太阳高度角的增加而增加(图1)。赤道地区光照强度的变化速率最大,而暮曙光最短;暮曙光随着纬度的增加(向两极)而增加(图2)。
随着地球自转速度的减慢,日光周期逐渐增加,变化速率约为0.002秒/世纪[113]。也就是说,如果一个物种的平均寿命五百万到一千万年[70],那么日长从该物种出现到灭绝只增加了约100-200秒,因此日光周期是一个非常可靠的因素。
自然光的光谱也会在一天中变化,特别是在黄昏时。当太阳从地平线以上的低处落到地平线以下时,在无月的夜晚,晴朗的天空会从白色变为微红,再变为蓝色,最后又变回微红;红光与远红光的比例也会在黄昏时期发生变化[56]。月光的影响有限,直到傍晚的时候,月相和高度才会显著地影响物体的颜色和亮度[79]。
1.2 季节光周期
地球的轨道运动和地轴相对于太阳的倾斜造成了季节性昼长、光照强度和暮曙光时长的变化,变化情况与纬度相关。虽然在不同的地区,每年的相对黑暗时间都约为4400小时(包括黄昏、月光和星光),但在高纬度地区,这种黑暗却集中在冬天(图1),而在低纬度地区全年均匀分布。低纬度地区黄昏时长全年基本保持不变,而中纬度地区全年变化较大(图2)。
每个图像代表在英国埃克塞特大学彭林校区气象站所记录的全天照度。在黄昏和黎明时段冬季月份的照度水平比夏季月份衰减和增加的更缓慢,日间照度差异也更显著。
1.3 月光周期
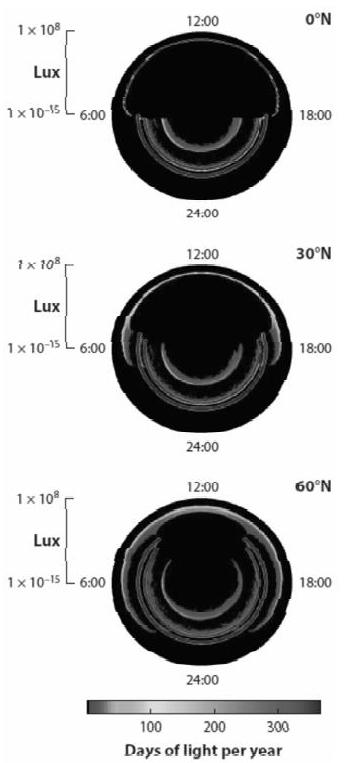
根据月球绕地球的运行轨道(周期29.5天),月球经过地球同一点所需的时间(24.8小时),以及由此反射产生的太阳光(月光)的强度和模式,所以夜间的光环境在不同时间尺度上受太阳光的昼夜和年周期变化的影响(图3)。与白天太阳光强度变化的数量级类似,夜间月光的光照强度变化幅度也跨越了三个数量级,虽然白天的光照强度明显高得多[60]。月光强度也受一年中的时间和纬度的影响;相较夏季,冬季月亮位于地平线之上更高的位置,因而夜晚更加明亮[60]。
日光周期、季节光周期和月光周期对地面接收的光照强度的影响受很多因素调控,而这些因素通常是多变且不可预测的,比如当地地形,生境和天气(特别是云层),另外间歇性的自然光源也有较小程度的影响(如雷击和火灾)[47]。然而,日光周期、季节光周期和月光周期的幅度和频率很大程度上是可预测的波动。因此在很多情况下,相比于潜在的其它因素(如,温度、降雨量、资源可用性),这些周期作为生物活动发生的线索更可靠。
一天内的时间用旋转轴表示,勒克司在径向轴上用对数值表示。颜色越红,一年中达到特定勒克斯水平的天数越多。在北纬0°,一年中太阳每天的光照强度没有变化,暮曙光很短,并且产生时间和持续时间一致。由于月亮的周期为29.5天,月光光照强度变化更大,但全月和新月的光照强度相对一致。随着纬度的增加,日光和月光达到了最大光照强度,暮曙光的产生时间和持续时间在年周期内变动更大,导致数据范围更广。在0.2-0.5lx和20-40lx的兩条带表明,人工白昼和街道照明可能掩盖了大部分月光,干扰了黎明开始和黄昏结束的判断。
2 人工照明对自然光周期的干扰
在夜间环境中引入人工照明,明显破坏了自然光周期。人工照明有两种形式。一种是来自光源的直接照度(图4)。通常发生在极为靠近光源的地方,如10-40lx路灯下地面的照度,在几米外通常会下降到1lx以下。(正常情况下地面照度要高得多,并且延伸得更远,例如在体育和港口设施等处的投光照明,以及在一些发展中国家贫困社区中使用的公用灯架)。直接照度可以直接影响生物体所感受到的光,特别是当生物体不能移动或移动性受限时,或能够在生物体移动的光影中产生空间异质性。来自人造光源的直接照度可以传播更长距离,特别是在水平面发射时,例如车辆前照灯;一辆普通的家用轿车发出的光可以达到10,000lx,距离50m处达25lx,距离100m时仍超过1lx[8]。由于光源的时空异质性,直接照度的整体范围很难测定。利用卫星传感器检测的数据估计光辐射(可能主要来自向上无屏蔽或屏蔽不良的路灯和反射光辐射)约为全球陆地面积的11%[47],但是这样的辐射只是光辐射的一部分,可能会低估地面的直接照度。
自然状态月空亮度模型(a)与英国普利茅斯市上空观测到的月空亮度(c)以及城市外的8.6公里处沿海地区(b)的对比。以上数据于2011年10月1日至2012年7月31日使用天空光度计(SQM-DL模型)进行测量[27]。
人工照明的另一种形式是人工白昼,主要是由向上发射的人造光在大气中被水、尘埃和气体分子散射造成的。能被积雪放大的地面反射的人造光也是形成人工白昼的原因[39]。人工白昼可以将夜空的背景亮度提高到0.2 -0.5 lx[37,59],与暮曙光和月光相当并且会阻碍人类对星空的观测(83%的人口生活在光污染的天空下)[39]。云层反射会显著地放大城市上空的人工白昼,同时降低更偏远地区的自然天空亮度[61]。城市人工白昼的放大程度随着云底的降低而增加,并且在可见光谱的红光端更加明显[2]。人工白昼在空间上必然比直接照明延伸范围更大。特别是有以相对较小角度发射向地平线的光源,人工白昼可以从光源延伸到数百公里之外[67]。据估计,全球约有23%的陆地经历人工白昼[39]。
一直以来,夜间人工照明的发射光谱和自然光(无论是太阳光还是月光)的发射光谱有很大差异。虽然全球范围内主流的照明类型因地而异,但是人造光源发射的光通常仅包括非常狭窄的波长范围,例如低压钠灯发出的光(在589.3nm处有一个窄峰)。然而,随着发光二极管(LED)技术越来越普及,虽然同阳光仍有明显差异,或就植被受到的光辐射而言,经常发生红移,发射光的光谱变得更宽(白色LED路灯波长通常在400和700 nm之间,在蓝绿区达到峰值)[8,38]。
2.1 日光周期的干扰
虽然具体人工照明变化取决于灯的种类和使用时间,但是人工照明往往在黄昏时开启,缩短了被照明区域的黑暗时间[97]。因此,夜间人工照明照亮了周边栖息地,延长了日长,但是光照并不会把夜晚变为白昼。人工照明对自然日长规律的干扰取决于国家或地区的城市照明政策[10]。虽然在一些地区商业、市政和工业用光会随夜深而关闭(车辆前照灯的影响同样下降),但是在其他地区,街道照明持续到黎明。最终导致城市地区整个夜晚的天空可变得比满月时更亮,随着夜深,夜空因人工光源的关闭而变暗;而附近的农村地区,随着月亮升起天空会变亮,月亮落下天空会变暗[61]。
由于夜间人工照明条件已知,人们认为估算不能移动的生物,在当地变化的日光周期(以及季节和月光周期)中的感光水平相对容易,当然被遮挡的以及用某种方式保护自己免受光照的生物体除外(如深层植被或地下)。相比之下,会移动的生物个体的感光水平可能更为复杂,特别是考虑到可能的趋光或避光行为。移动生物体感知到的夜间人工照明强度的直接测量方法仍然非常有限,但是利用安装在动物身上的光记录仪,研究人员发现,与生活在农村森林的欧亚乌鸫(Turdus merula)相比,受照度的增加的城市和工业区的欧亚乌鸫光照活动时间延长[32]。对于穿梭于路灯下的光斑夜行动物,能感受的光照强度会由三个数量级的自然光强度增加到约五个数量级。对于栖息在路边的夜行动物,车前灯使它们感受的光照强度将增加到约六个数量级。
2.2 对季节光周期的干扰
因为夜间照明是为了给人类照亮黑夜,所以人工夜间照明通常会随着季节和纬度变化而变化。夜间有了人工照明,黎明似乎开始得更早,而黄昏结束得更晚,甚至在某些情况下,暮曙光可能充斥整个夜晚。从中纬度到高纬度地区,日长的变化提供了重要的可以预测气候条件和资源可用性的生物线索。因为低纬度地区日长的变化不显著,物候学与降雨和温度变化密切相关,所以与低纬度地区相比,人工照明对高纬度地区的生物物候学的干扰更大。假设路灯照明下日长的变化仍然可以预测,生物体仍然能够在人工照明区域进行合适的季节性生物活动;然而,这些活动还是会因日长变化的干扰而改变[33,86]。由于一个群落里的不同个体受到的影响不同,多种生物的生活史之间,也可能发生错配[52]。高纬度地区冬季夜晚更长,意味着人工照明对冬季活动物种适应性的影响,比对那些仅在早春至深秋季节活跃或出现的物种的影响更大,更极端的是,一些本应24小时处于黑暗中的北极定居点也局部出现了光亮。
2.3 对月光周期的干扰
与日光周期和季节光周期的昼长相比,夜间人工照明更容易掩盖月光周期的环境线索。这可能只是因为月光强度比日光强度要弱得多。实际上,仅仅人工白昼就足以掩盖自然状态下月夜天空亮度的季节性变化,让生物在夜晚接受到了相当于自然条件下满月时才能感受到的强度光照(图3)[27]。因此,生物的月光周期在一些受夜间人工照明更直接影响的地区不明显。另外,人工光源的光谱会干扰生物感知月光影响的暮曙光光谱的能力,而这是如海洋无脊椎动物产卵活动等特定生物活动发生的潜在重要线索[103]。
3 对日常生物活动的影响
对于夜间人工照明对日常(或其它周期)野外生物活动影响的理解大部分仍然停留在表象。也就是说,虽然很多生物活动的节律变化被记录下来,但是最为重要的驱动这些变化的干扰自然光周期的因素目前尚不清楚。人们一致认为,光照强度是关键的驱动因素,也是已知的自然光条件下重要的节律调节因子。然而,考虑到人造光会同时干扰光照强度和光谱,所以应该还有光照强度之外的影响因素。值得注意的是,尽管人们早已熟知哺乳动物的生物钟受光强度变化的影响,但最近发现光谱变化的影响也很重要[115]。
目前人们在野生生物感知夜间人工照明程度和光谱方面,对剂量-响应关系以及生物节律是如何受光强和光谱的影响的认识也所知甚少。已知的剂量-响应研究通常,只关注实验条件下,不同人工光源造成的影响,而不是真正的野生生物生存的环境中的特定类型的光源条件[18,30]。
人工照明对日光周期、季节光周期和月光周期的影响很大程度上依赖环境。如在极端条件下,由于温度的限制,将人工照明引入高纬度冬季全天24小时黑暗地区所造成的影响,并不同于通常在夏季才活跃的生物。虽然在野生生物中还有待探索,夜间人工照明的影响也可能受到生物此前感知光照方式的影响[5]。
3.1 鸟鸣
夜间人工照明对温带地区鸟鸣节律的影响一直是人们关注和研究的主题。有证据表明在有夜间人工照明的地区,鸟类清晨和傍晚鸣叫的时间会提前,这对繁殖有影响[25,57,72]。(也有证据表明,在有夜间人工照明下鸟鸣时节也会提前[24]。)由于热带地区昼长的季节变化有限,光照强度引发鸟鸣大大减少。事实上,在热带的研究并没有发现夜间人工照明对鸟鸣节律有任何显著影响[35]。
3.2 觅食
夜间人工照明已被证明可延长一些昼出动物的觅食时间并会减少一些夜行动物的觅食时间[4,14,40,43,62,64,76,81,93,118]。夜间人工照明会改变生物的觅食方式,捕食者可能利用光照捕获猎物,猎物也可能利用光照躲避追捕[13,19, 36,51,114]。人工照明也可能吸引猎物聚集,反过来又会增加一些捕食者的觅食机会[20,73,99]。然而,不同物种做出反应仍很难预测,即便是同类也会表现出完全不同的行为[88]。
人们对夜间人工照明和觅食的剂量-响应关系还知之甚少,虽然最近有研究表明大山雀(Parus major)的活动与光强度有很密切的联系,即使光强增加很少,大山雀的夜间活动也会增加[31]。同样,有少量研究比较了夜间人工照明的不同光谱对觅食的影响,发现影响差异很大[89,101]。
目前还不清楚夜间人工照明对觅食节律的影响与整体捕食率、猎物种群规模是否有因果关系。虽然有一项研究发现其对鳞翅目幼虫的捕食率没有影响,但其他研究人员认为,现代广谱照明技术的广泛使用可能会导致夜行和昼行动物之间对食物资源的竞争,以及改变主要陆生动物间的物种平衡[28,48,68]。
3.3 昼夜活动
毫无疑问,夜间人工照明与觅食相关,同时夜间人工照明也改变了许多生物的日常活动时间。最明显的例子可能是,在水生系统中浮游生物的昼夜垂直迁移对细微的自然光照变化非常敏感,因此会受到夜间人工照明的影响[11,74]。考虑到水生系统中这两种光照的水平,以及浮游生物可能是全球最大的生物质和碳元素同步循环系统,夜间人工照明对浮游生物昼夜活动的干扰可能是所有夜间人工照明影响中最广泛的[29]。
许多其他生物的昼夜活动也受到避光行为的影响,这表明通过减少和破坏天然黑暗栖息地,人工照明干扰了夜间避光生物获取生存资源[49]。许多避光生物(如木虱、千足虫、蜈蚣、环节动物)对分解和营养循环十分重要,人工照明影响它们正常的活动模式可能会导致土壤质量下降。
当然,基于特定光谱在吸引物种上有明显的效果,所以夜间很多物种会直接(如许多蛾类)或间接(如一些蝙蝠物种会捕食在灯周围的蛾类)靠近人工光源[65,90,96,109]。包括一些人类疾病的重要载体,如南美锥虫病的传染媒介——二分锥蝽(Triatoma dimidiata)。该物种更喜欢出没于路灯附近的房屋,更易受短波的吸引[78]。
由于人工照明對生物活动有影响,光源在单个建筑以及整体上的分布对物种的分布模式和丰度影响显著[6,16]。实际上,市区人工光源密布的市区特别明显,既可能是夜晚生物穿梭的走廊,也可能是夜晚生物穿行的枷锁。
关于夜间人工照明的强度对生物进化的影响的研究仍然很少[102]。然而有研究显示,幼虫时期即在高亮度人工照明地区饲养的卫矛巢蛾(Yponomeuta cagnagella)种群与那些来自天然光照环境的种群相比,趋光行为会显著减少[1]。这种进化的改变意味着,动物在自然光照条件下有助于生存的一些天性,如趋光性,由于人工照明的危害,在市区的昆虫群落中已经退化。
3.4 睡眠
人工夜间照明会影响动物的睡眠模式。虽然目前观察到的这些作用的生理机制(特别是短波长光能够抑制褪黑激素产生)很大程度上适用于哺乳动物以及其它类型的脊椎动物,但是迄今为止,大多数研究还只是在人类中进行[21,117]。一项针对雌性大山雀(P.major)的实验表明,在人工夜间照明下,她们往往入睡晚,起床早,睡眠时间较少[82]。睡眠模式的破坏对生物健康有显著影响,因此探讨在广泛的生物类群中,这些影响的程度是有价值的。
