甘蔗响应梢腐病菌侵染的蛋白质组学分析
2019-06-20王泽平林善海梁强李长宁宋修鹏刘璐李毅杰
王泽平 林善海 梁强 李长宁 宋修鹏 刘璐 李毅杰
摘 要 本文旨在解析甘蔗响应梢腐病菌侵染过程中的生理生化机制,为甘蔗抗梢腐病育种及病害防治提供理论指导和科学参考。以我国甘蔗梢腐病主要致病菌Fusarium verticillioides孢子悬浮液为病原,以高抗梢腐病品种YT94/128和高感梢腐病品种GT37为宿主材料,在温室条件下进行针刺法接种,提取病情指数最严重时期,即接种后第14天的甘蔗葉片蛋白质进行iTRAQ定量表达分析。结果显示,从GT37中成功鉴定3707个蛋白,获得542个差异表达蛋白,其中上调表达187个,下调表达355个;从YT94/128中成功鉴定到3068个蛋白,获得差异蛋白449个,其中上调表达191个,下调表达258个。这说明遗传背景不同的甘蔗品种在蛋白质组成上有很大差异,推测这是不同甘蔗品种间抗梢腐病性差异的重要分子基础。
关键词 甘蔗;梢腐病;轮枝镰孢菌;蛋白质组学;侵染
中图分类号 S566.1 文献标识码 A
Abstract This study was to provide a theoretical guidance and scientific reference for sugarcane resistance breeding and disease prevention by analyzing the physiological and biochemical mechanism in the infection process of sugarcane response to pokkah boeng disease. With spore suspension of Fusarium verticillioides (the mainly pathogenic fungus of pokkah boeng disease in China) as the pathogeny and YT94/128 (HR), GT37 (HS) as the tested materials by syringe inoculation under a greenhouse, and the samples were collected at the 14th post-inoculation with the highest disease index to carry out quantitative expression analysis by isobaric tags for relative and absolute quantitation (iTRAQ). The results showed a total of 3707 proteins were successfully identified from GT37, and 542 differentially expressed proteins (DEPs) were obtained, of which 187 were up-regulated and 355 were down-regulated. Meanwhile, a total of 3068 proteins were successfully identified from YT94/128, and 449 DEPs were obtained, of which 191 were up-regulated and 258 were down-regulated. In a word, there are significant differences in the protein composition of sugarcane varieties with different genetic backgrounds, and it is speculated that this is the important molecular basis for the difference of resistance on pokkah boeng disease between different sugarcane varieties.
Keywords sugarcane; pokkah boeng disease; Fusarium verticillioides; proteomic; infection
DOI 10.3969/j.issn.1000-2561.2019.05.015
甘蔗梢腐病(pokkah boeng disease)是由镰刀菌(Gibberella fujikuroi)引起的,在我国蔗区其主要病原为Fusarium verticillioides[1]。该病无季节性流行,导致甘蔗表现出不同程度受害,已给蔗糖业可持续性生产造成严重影响[2]。
在生物或非生物胁迫条件下,甘蔗蛋白的含量和种类会发生变化,其表达丰度或增或减,或产生新的蛋白。乙醇脱氧酶、类异黄酮还原酶、咖啡酸3-O-转移酶、谷胱甘肽转移酶、抗坏血酸过氧化物酶、USP家族蛋白以及胱硫醚β-合酶参与甘蔗防卫宿根矮化病菌反应[3];甘蔗可溶性蛋白在盐胁迫下的表达存在显著差异[4];机械损伤诱导蛋白SUGARWIN2 在赤腐病、凤梨病病原菌及酵母菌诱导下表达迥异[5];甘蔗拓补异构酶,乙烯不敏感因子以及4次穿膜蛋白3个基因参与防御黑穗病菌侵染[6];蔗芽抗病性涉及活性氧代谢途径和苯丙烷代谢途径关键酶活性的变化[7];病程相关蛋白PR-1和β-1,3葡聚糖酶等被诱导上调表达[8];甘蔗可能通过细胞色素C氧化酶基因的诱导,促使植保素合成增多,以此抵抗或抑制病原菌的胁迫[9]。甘蔗抗梢腐病菌F. verticillioides侵染研究表明二者相互作用机制涉及RNA、可溶性蛋白及氨基酸含量的精细调节[10]。
同位素标记相对和绝对定量技术(Isobaric tag for relative and absolute quantitation, iTRAQ)作为近年来发展的蛋白定量技术,其已成功应用于解析甘蔗防御外界环境胁迫涉及的多系统、多水平代谢进程[11]。为此,本文拟在上述研究基础上,利用iTRAQ技术监测梢腐病菌诱导的甘蔗叶片蛋白的表达水平变化,以获得甘蔗响应梢腐病菌侵染的差异蛋白表达谱,为解析甘蔗抗梢腐病生理生化机制提供全景扫描图,进而为发掘病程相关蛋白和抗病基因提供理论指导和科学依据。
1 材料与方法
1.1 材料
室内条件下种植高抗梢腐病品种YT94/128和高感品种GT37,待其5~6叶时,选取长势一致植株利用针刺法[12]接种病原菌F. verticillioides,并遮荫保湿,保持大棚温度20~35 ℃,湿度80.00%~85.00%。于病情指数最严重阶段即接种后第14天采集甘蔗+1叶(接种病原菌和清水的2个甘蔗品种,各3次生物学重复,共计12个样品)经液氮速冻,保存于80 ℃冰箱备用。
1.2 方法
1.2.1 蛋白质提取及测定 按照每106个细胞加约400 μL 裂解液的比例(7 mol/L尿素,2 mol/L硫脲,0.1% PMSF,65 mmol/L DTT),在样品中加入合适的裂解液,冰浴超声(超声5 s停10 s,超声3~5 min);冰上静置40 min;14 000 r/min 4 ℃离心30 min,取上清。参考苏亚春[8]的方法进行蛋白质定量和检测。
1.2.2 蛋白质电泳、烷基化及酶解 SDS-PAGE电泳:取出需要的样本量加入终浓度为10 mmol/L DTT,在56 ℃下反应30 min后,加入终浓度20 mmol/L IAA室温下避光反应30 min;每管各加入预冷的丙酮(丙酮∶样品体积比=5∶1),20 ℃沉淀2 h;12 000 r/min,4 ℃,离心20 min,取沉淀;加入含1 mol/L尿素的TEAB溶解液(dissolution buffer)20 μL,充分混匀溶解样品;按照质量比1∶50(酶∶蛋白)加入Trypsin,在37 ℃酶解15 h;酶解液加入终浓度0.5%的TFA终止酶解,浓缩冻干。
1.2.3 iTRAQ标记及质谱分析 胰蛋白酶消化后,用真空离心泵抽干肽段;用0.5 mol/L TEAB复溶肽段,按照魏春燕[11]方法进行iTRAQ(Applied Biosystems, Foster City, CA, USA)标记。
高pH反相液相色谱法(RPLC)第一维分离:2.1 mm×150 mm XBridge BEH300(Waters公司,USA);色谱仪器:Waters UPLC;A相:水(氨水、甲酸调至pH 10);B相:100%CAN;紫外检测波长:214~280 nm;流速:200 μL/min;梯度:60 min。根据峰型和时间共收取20个馏分,真空离心浓缩后,用50 μL RPLC A相溶解,进行第二维分析。采用Triple TOF 5600(Applied Biosystem,USA)质谱仪进行LC-TMS分析,采集软件:Analyst TF(ABI,USA);反相柱:ZORBAX 300SB C18 色谱柱(5 ?m,300 A,0.1 mm×150 mm,microm,USA);色谱仪器:Eksigent 1D plus;色谱分离90 min;A: 5%CAN,0.1%甲酸;B:98% CAN,0.1%甲酸;流速:300 μL/min。
1.2.4 蛋白质鉴定及生物信息学分析 应用Mascot 2.3.02 软件,在西南大学家蚕基因组数据库SilkDB(http://silkworm.swu.edu.cn/silkdb/doc/ download.html)中进行蛋白质的鉴定和定量。最终选择肽段数≥2,表达水平差异倍数>1.0(上调)或<1.0(下调),且P<0.05的蛋白质为显著差异表达蛋白。选择欧洲生物信息研究所(European Bioinformatics Institute,EMBL-EBI)维护的QuickGO(http://www.ebi.ac.uk/QuickGO/)注释工具对差异蛋白质进行基因功能聚类GO分析,把所有差异表达蛋白质向Gene Ontology 數据库(http://www.geneontology.org/)的各个条目映射,计算分布在每个条目的蛋白质数目,然后应用超几何检验,找出与所有蛋白质背景相比在差异蛋白质中显著富集的GO条目。Pathway通路显著性富集分析方法与GO功能富集分析方法相同,以KEGG Pathway为单位,应用超几何检验,找出与所有鉴定到的蛋白质背景相比,在差异蛋白质中显著性富集的Pathway。
2 结果与分析
2.1 蛋白质组的质谱鉴定结果
接种梢腐病菌后,GT37与对照相比有542个差异表达蛋白,其中上调表达187个,下调表达355个。YT94/128较之对照获得差异蛋白449个,其中上调表达191个,下调表达258个。接种清水后,YT94/128较之GT37有131个差异蛋白下调表达,87个蛋白上调表达,而在接种病原菌后,YT94/128较之GT37有44个差异蛋白下调表达,151个蛋白上调表达。
2.2 差异表达蛋白质的GO功能注释
GT37中注释到3707个蛋白,其中生物学过程(biological process)中蛋白数目最多的是代谢进程(metabolic process),为322个;分子功能(molecular function)中蛋白数目最多的则是催化活性(catalytic activity),共计299个,细胞成分(cellular component)中蛋白数目最多的是细胞组分(cell part)和细胞(cell),二者均为419个。YT94/128中注释到3068个蛋白,其中生物学过程中蛋白数目最多的是细胞进程,为285个,分子功能中蛋白数目最多的是催化活性,为237个,细胞成分中蛋白数目最多的是细胞组分和细胞,二者均为345个。
2.3 差异表达蛋白质的KEGG通路富集分析
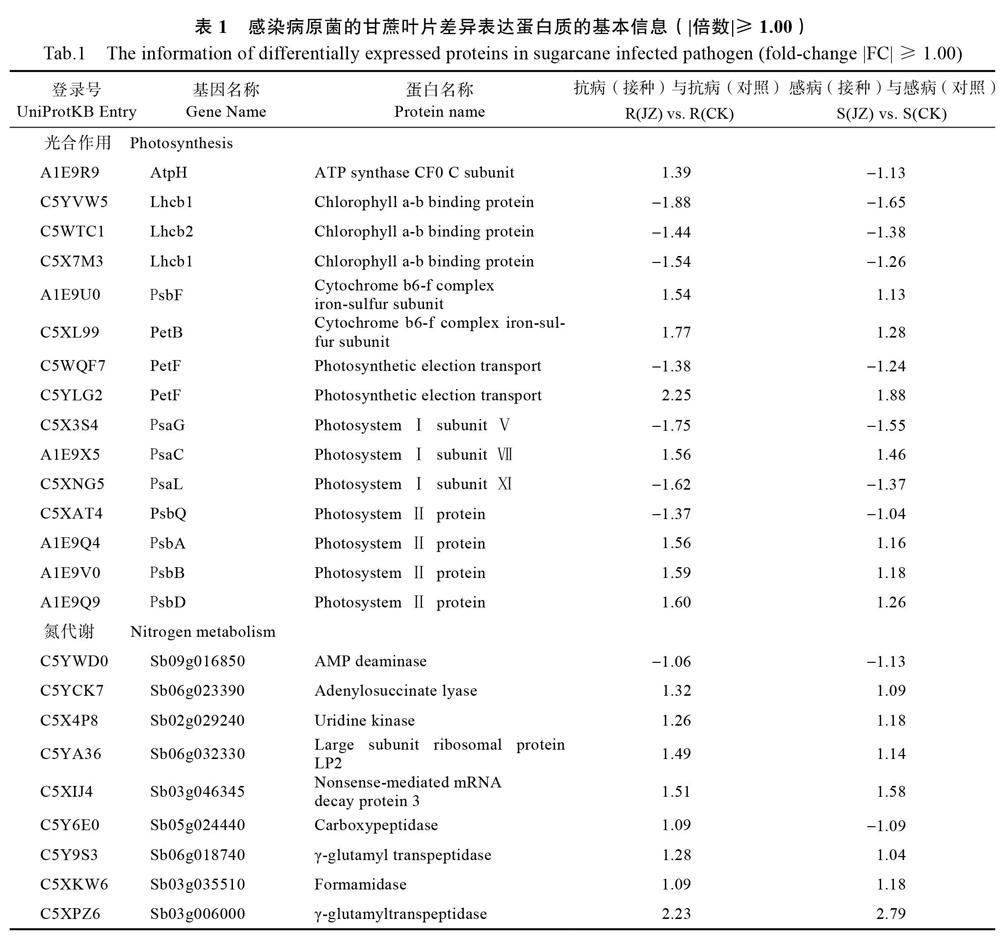
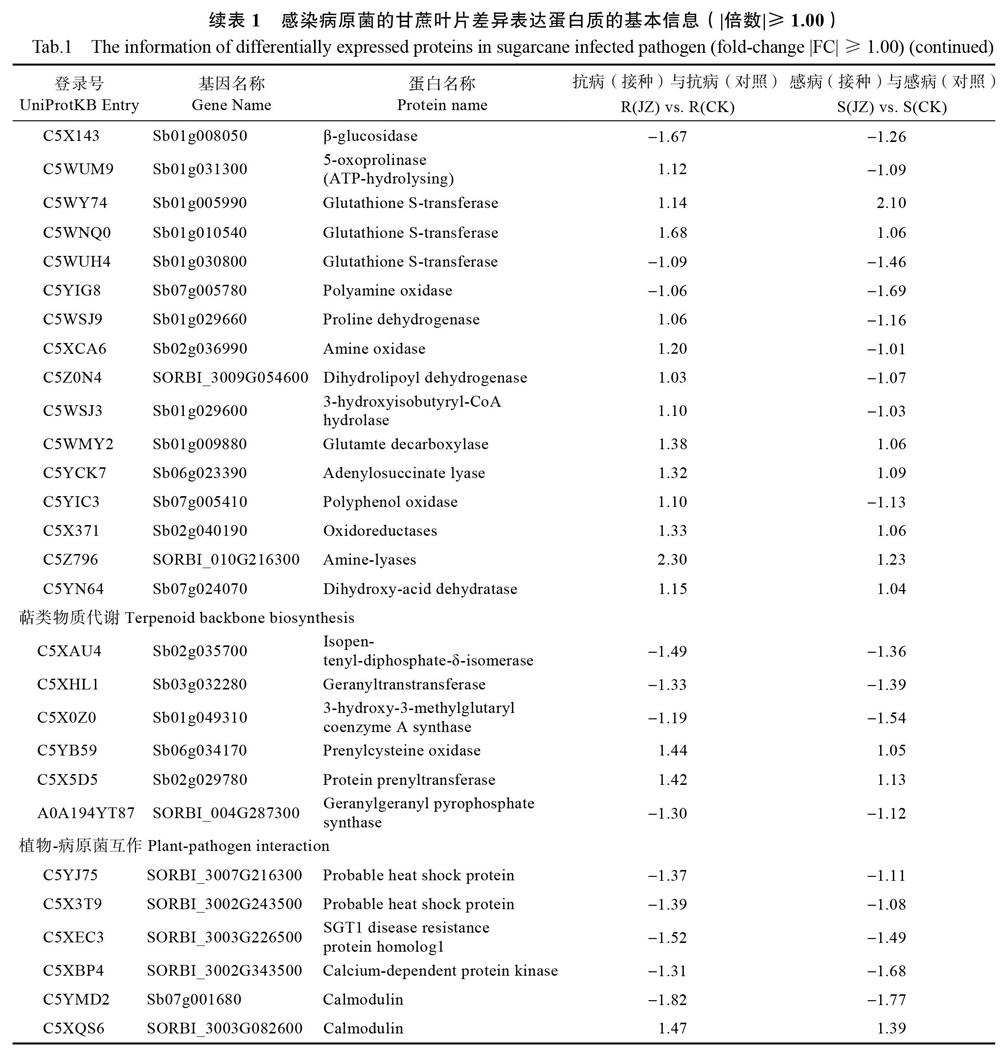
GT37中主要富集的通路有光合生物体碳固定(carbon fixation in photosynthetic organisms)、碳代谢(carbon metabolism)、戊糖磷酸途径(pentose phosphate pathway)、光合作用(photosynthesis)、氨基酸生物合成(biosynthesis of amino acids)、丙酮酸盐代谢(pyruvate metabolism)、亚油酸代谢(linoleic acid metabolism)、光合作用捕光蛋白(photosynthesis/antenna proteins)、乙醛酸和二羧酸代谢(glyoxylate and dicarboxylate metabolism)和代谢途径(metabolic pathways)。YT94/128中主要富集的通路有亚油酸代谢、乙醛酸和二羧酸代谢、光合作用、光合作用捕光蛋白、苯丙烷类生物合成、碳代谢、谷胱甘肽代谢、萜类物质代谢、次生代谢物质合成和丙酮酸盐代谢。将通路富集信息与定量分析结果整合见表1,得到部分重要差异蛋白质的基本信息,其中主要包括15个光合作用相关蛋白、25个氮代谢蛋白、6个萜类物质代谢蛋白和6个植物-病原菌互作蛋白。
CK、JZ分别表示采用清水和孢子悬浮液接种第14天的样品。
3 讨论
本文基于iTRAQ组学技术构建了梢腐病菌侵染不同甘蔗材料的差异蛋白表达谱。代谢通路分析发现叶绿素a-b结合蛋白(Lhcb1-3)作为光合作用部位关键性成分在光捕获和光调节方面发挥重要作用[13]。此类蛋白下调表达,说明在病原菌诱导的衰老和死亡过程中控制PSⅡ-捕光复合体(PSⅡ-LHCⅡ)动力的类囊体结构蛋白(THF1)受到负调控。三磷酸腺苷合成酶(ATP synthase,α、β、γ)作为病原菌侵染潜在受体在受到胁迫后上调表达,暗示梢腐病菌可通过影响宿主甘蔗ATP 合成酶的活性来扰乱其正常能量代谢。细胞色素b6f蛋白复合体(PetB和PetC)在PSⅠ和 PSⅡ反应中心的产氧光合作用和跨膜电化学质子梯度中为ATP合酶提供电路连接[14],各类植物中,细胞核编码因子在Cyt b6f复合体转录过程中是必不可少的,诸如玉米中的CRP1蛋白、衣藻属的MCA1蛋白、以及拟南芥的PGR3、HCF152、HCF153蛋白。在光保护机制诱导下,过氧化物酶能协助清除过氧化氢,此类蛋白上调表达说明甘蔗在受到病原菌胁迫后,其叶片氧化还原反应速度显著加快。PSⅠ(由PsaC、PsaG和PsaL组成)参与电子传递和状态转换以维持高效光合作用,其大部分组合因子在蓝藻细菌、真核藻类和CK、JZ分别表示采用清水和孢子悬浮液接种第14天的样品。
高等植物中具有高度保守性[15],除C5XAT4以外,大部分显著上调表达,据此推断C5XNG5、C5X3S4、A1E9X5以及PSⅠ的其他组成部分可能共同产生了一组聚合体,它们能确保对病原体侵染的快速响应和光合效率的正常进行。PSⅡ周围的细胞色素蛋白参与环式电子传递调节,且对外界环境诱导有保护效应。由此可见,光合作用机制自动响应促使甘蔗调控适应外界胁迫,避免真菌对正常代谢途径的进一步侵扰,或使得甘蔗植株在受到较轻感染后能尽快恢复生长。相反,感病基因型甘蔗却无法产生足够的适应性。
甘蔗叶片氮代谢通路积极响应梢腐病菌的侵染[10]。C5Z0N4作为二氢硫辛酸脱氢酶以及C5WSJ3作为3-羟基异丁酰辅酶A水解酶参与半胱氨酸和蛋氨酸代谢途径,而半胱氨酸是硫酸盐同化作用的主要初级产物,硫素缺乏可导致叶片褪绿黄化、心叶伸长迟缓及花青素和大量细胞内含物的积累,这与氮素供应受限或紊乱所致症状极其相似,说明抗感差异材料中利用此类蛋白机制是迥然相反的。在β-丙氨酸代谢过程中,β-丙氨酸可能通过胺氧化酶(C5XCA6)、多胺氧化酶(C5YIG8)或谷氨酸脱羧酶(C5WMY2)进行氧化脱氨基作用形成一些半醛,然后经过重组参与丙酮酸代谢进程,反过来又通过胺化作用产生β-丙氨酸。胺氧化酶(C5XCA6)和脯氨酸脱氢酶(C5WSJ9)在YT94/128中上调表达,在GT37中下调表达,表明抗病性更强的甘蔗材料可能具有更活跃的胺代谢分解。C5XPZ6作为γ-谷氨酰转移酶和谷胱甘肽水解酶以及C5WY74、C5WNQ0作为谷胱甘肽S-转移酶在感抗植物中上调表达,表明它们在谷氨酸代谢中都扮演了非常重要角色。C5XPZ6又作为白三烯C4水解酶发挥重要作用,这说明该酶发挥的生理生化功能与甘蔗获得系统性抗性有关。系统性抗性典型性特征表现为防御广谱抗菌,并独立于微生物或化学诱导物用于初级诱导。因此,进一步鉴定花生四烯酸是否是为系统性抗性真正诱导者,或在甘蔗中体内流动存在发挥作用是非常有必要的。
基于3-氨基丙腈具有抗病毒活性,因此氨腈代谢通路也被认为与甘蔗抗梢腐病机制相关。C5Z796,作为一种胺裂解酶,在感抗材料中上调表达,尤其是在YT94/128中显著上调,参与了赖氨酸代谢过程中L-天冬氨酸4-半醛和(2S, 4S)-4-羟基-2, 3, 4, 5-四氢二吡啶的反应。当细胞碳水化合物供应被耗尽时,其可作为另一种呼吸底物发挥重要作用,而二氢吡啶二羧酸合成酶(DHDPS)保持正常调节水平,赖氨酸是不会增长的[16]。在同批接种样品中,感抗材料中不仅赖氨酸含量较对照显著增长[10],参与赖氨酸代谢进程并起主要催化还原活性的C5Z796蛋白也显著上调表达。因此,C5Z796蛋白被认为是甘蔗与病原菌互作的一种特征性蛋白。
萜类化合物由于其挥发性、风味/香气和毒性等特性,在植物防御中发挥重要作用 [17]。通路富集分析表明萜类蛋白C5XAU4作为焦磷酸-δ-异构酶参与类异戊二烯及甾醇生物合成,C5XHL1作为催化酶参与香叶酸合成,C5YB59參与脱落酸信号转导、法内二磷酸及异戊烯半胱氨酸的合成,C5X5D5参与脱落酸信号途径的负调控、蛋白的法尼基化修饰、香叶酰香叶酰化、分生组织的形成及水分丧失响应过程,C5X0Z0将乙酰辅酶a与乙酰辅酶a结合形成β-羟基-β-甲戊二酸单酰辅酶a。这些反应都是在一系列萜烯合成酶催化下进行,中间合成产物有许多是次生代谢产物,它们可以保护甘蔗不受梢腐病菌的侵害。在受感染的植物中,萜烯的生物合成被抑制,降低了植物抗性,增强了病毒的传播性能[18]。基于此,针对萜类化合物是否能作为毒素抑制梢腐病菌的生殖和繁育、是否存在特定的萜烯化合物使梢腐病菌产生排斥反应影响对不同遗传背景甘蔗的选择行为以及是否存在特定的萜烯化合物通过吸引天敌(内生菌)间接防御梢腐病菌的侵染这一系列问题开展进一步探讨是非常有必要的。
植物寄主抗病性与其合成的多肽有关。蛋白C5YJ75和C5X3T9具有ATP酶活性,可能与RAR1结为协同分子伴侣积极参与RPM1介导的抗性机制。前人已通过大麦白粉病研究发现了RAR1蛋白在抗病信号中的作用[19],在一些属于NBS-LRR家族的基因(12, 36-38)中,也发现了同样的作用[20]。此外,Hsp90可以稳定RPM1并保护它不受Sgt1介导的降解,在无热休克情况下通过主动抑制热休克转录因子功能,对热诱导基因进行负调控[21]。C5XEC3被注释为Sgt1 disease resistance protein,暗示其可能参与植物与病原菌的互作进程。所有外界环境变化都可导致第二信使钙(Ca2+)信号的产生[22],钙依赖性蛋白激酶(CDPKs),代表潜在的Ca2+解码器以转化发育和环境应激信号。蛋白C5XBP4通过参与ATP的结合、催化钙依赖性蛋白丝氨酸/苏氨酸激酶的活性以及与Ca2+和钙调蛋白的结合积极参与脱落酸信号转导、胞内信号转导、肽基丝氨酸磷酸化以及蛋白自身磷酸化等生理过程,表明其可能在快速响应病原菌胁迫,且在植物激素调节适应性方面发挥重要作用。前人通过对大豆钙调蛋白(CaMs)的过度表达研究,证明了CaM参与植物防御反应的直接证据[23]。在转基因烟草和拟南芥中,SCaM4和SCaM5的表达会导致自发损伤,增加PR基因表达,增强对细菌、真菌和病毒病原体的抵抗力[24]。有趣的是,沉默烟草中的一个CaM(Nb-CaM1),抑制烟草细胞中烟草花叶病毒p50诱导的HR,而不是Cf9-Avr9或Pto-AvrPto和pst dc3000诱导的细胞死亡,说明CaM可以对不同的病原体产生特异性[25]。这表明CaMs在植物防御中起着至关重要的作用,也暗示了钙调蛋白C5YMD2以及C5XQS6在甘蔗材料中有助于防御梢腐病菌的进一步侵染。
综上所述,遗传背景不同的甘蔗品种在蛋白质组成上有很大差异,这可能是不同甘蔗品种间抗梢腐病性差异的重要分子基础。而光合作用电子传递蛋白C5YLG2、γ-谷氨酰转移酶C5XPZ6、胺氧化酶C5Z796以及钙调蛋白C5YMD2作为高丰度差异表达蛋白,应可视为潜在重要抗病相关蛋白,其表达特性和功能特征有必要进一步深入研究。
参考文献
Lin Z Y, Xu S Q, Que Y X, et al. Species-specific detection and identification of Fusarium species complex, the causal agent of sugarcane pokkah boeng in China[J]. PLoS One, 2014: 9(8): e104195.
王泽平, 陈 奕, 孙海军, 等. 广西甘蔗主栽品种梢腐病田间抗性初步评价[J]. 热带作物学报, 2016(5): 952-957.
谢晓娜. 宿根矮化病菌侵染后甘蔗防御酶活性和蛋白质表达的变化[D]. 南宁: 广西大学, 2014.
Pacheco C M, Pestana-Calsa M C, Gozzo F C, et al. Differentially delayed root proteome responses to salt stress in sugar cane varieties[J]. Journal of Proteome Research, 2013, 12(12): 5681-5695.
Franco F P, Santiago A C, Henrique-Silva F, et al. The sugarcane defense protein SUGARWIN2 causes cell death in Colletotrichum falcatum but not in non-pathogenic fungi[J]. PLoS One , 2014, 9(3): e91159.
Huang N, Zhang Y Y, Xiao X H, et al. Identification of smut-responsive genes in sugarcane using cDNA-SRAP[J]. Genetics and Molecular Research, 2015, 14(2): 6808.
宋修鵬. 黑穗病菌侵染初期甘蔗幼苗光合生理变化及基因差异表达[D]. 南宁: 广西大学, 2014.
苏亚春. 甘蔗应答黑穗病菌侵染的转录组与蛋白组研究及抗性相关基因挖掘[D]. 福州: 福建农林大学, 2014.
阙友雄, 许莉萍, 张木清, 等. 甘蔗中一个NBS-LRR类基因的全长克隆与表达分析[J]. 作物学报, 2009, 35(6): 1161-1166.
王泽平, 李毅杰, 梁 强, 等. 不同甘蔗在梢腐病菌侵染下氮代谢相关指标的变化[J]. 植物生理学报, 2017, 53(11): 1963-1970.
魏春燕. 内生固氮菌DX120E与甘蔗互作的生理和分子生物学基础研究[D]. 南宁: 广西大学, 2016.
Wang Z P, Sun H J, Guo Q, et al. Artificial inoculation method of pokkah boeng disease of sugarcane and screening of resistant germplasm resources in subtropical China[J]. Sugar Tech, 2017, 19(3): 283-292.
Galka P, Santabarbara S, Khuong T T H, et al. Functional analyses of the plant photosystem I-light-harvesting complex II supercomplex reveal that light-harvesting complex II loosely
bound to photosystem II is a very efficient antenna for photosystem Ⅰ in state Ⅱ [J]. The Plant Cell, 2012, 24(7): 2963-2978.
Saif Hasan S, Yamashita E, Cramer W A. Transmembrane signaling and assembly of the cytochrome b6f-lipidic charge transfer complex[J]. Biochimica et Biophysica Acta, 2013 1827(11-12): 1295-1308.
Yang X F, Wang Y T, Chen S T, et al. PBR1 selectively controls biogenesis of photosynthetic complexes by modulating translation of the large chloroplast gene Ycf1 in Arabidopsis[J]. Cell Discovery, 2016, 2: 16003.
Amir R, Galili G. Regulation of lysine and threonine metabolism in plants[M]// Setlow J K. Genetic Engineering: Principles and Methods, vol 21. Boston: Springer, 1999: 57-77.
Chen F, Tholl D, DAuria J C, et al. Biosynthesis and emission of terpenoid volatiles from Arabidopsis flowers[J]. The Plant Cell, 2003, 15(2): 481-494.
Susi H, Barrès B, Vale P F, et al. Co-infection alters population dynamics of infectious disease[J]. Nature Communications, 2015, 6: 5975.
Zhou F, Kurth J, Wei F, et al. Cell-autonomous expression of barley Mla1 confers race-specific resistance to the powdery mildew fungus via a Rar1-independent signaling pathway[J]. The Plant Cell, 2001, 13(2):337-350.
Jo J, Venkatesh J, Han K, et al. Molecular mapping of PMR1, a novel locus conferring resistance to powdery mildew in pepper (Capsicum annuum)[J]. Frontiers in Plant Science, 2017, 8: 2090.
Blacklock K, Verkhivker G M. Allosteric regulation of the Hsp90 dynamics and stability by client recruiter cochaperones: protein structure network modeling[J]. PLoS One, 2014, 9(1): e86547.
Brandt B, Munemasa S, Wang C, et al. Calcium specificity signaling mechanisms in abscisic acid signal transduction in Arabidopsis guard cells[J] eLife, 2015(4): e03599.
Walton S D, Chakravarthy H, Shettigar V, et al. Divergent soybean calmodulins respond similarly to calcium transients: insight into differential target regulation[J]. Frontiers in Plant Science, 2017, 8: 208.
Reddy A S N, Ali G S, Celesnik H, et al. Coping with stresses: roles of calcium- and calcium/calmodulin-regulated gene expression[J]. The Plant Cell, 2011, 23(6): 2010-2032.
Zhu X H, Caplan J, Mamillapalli P, et al. Function of endoplasmic reticulum calcium ATPase in innate immunity-mediated programmed cell death[J]. The EMBO Journal, 2010, 29(5): 1007-1018.
