雷公山国家级自然保护区棉革菌属大型真菌形态描述及分子鉴定
2019-06-12张文泉罗国涛姚庆智
张文泉,罗国涛,姚庆智
(1.凯里学院,贵州 凯里 556011;2.内蒙古农业大学 生命科学学院,内蒙古 呼和浩特 010018)
棉革菌属Tomentella Pers.ex Pat 属担子菌门Basidiomycota 伞菌纲树种及水青冈Fagus longipetiolata Seem,桦木Betula 等阔叶树种形成外生Agaric-omycetes 革菌目Thelephorales 革菌科Thelephoraceae[1-2],常与马尾松Pinus massoniana Lamb,油松Pinus tabuliformis Carriere 等针叶菌根[3-4],与杜鹃Rhododendron simsii Planch,兰花Cymbidiumss 等草本或灌木形成内生菌根[5-7]。在森林生态系统中,菌根扮演着非常重要的角色,对其稳定性起着至关重要的作用,而对棉革菌属真菌研究发现,几乎棉革菌属的所有种均能形成菌根,因此,在森林生态系统中,棉革菌的作用不容忽视,具有重要意义[8]。
由于棉革菌属的子实体颜色以深色为主,多呈絮状或蛛网状平伏与腐木,枯枝或石头,土洞上紧贴生长,难以发现,因此国内外对于棉革菌属的研究多集中于棉革菌属所形成的外生菌根上。国内对棉革菌属的研究均为棉革菌属外生菌根研究:谢雪丹等对云南松幼苗上6 种Tomentella 外生菌根真菌形态进行报道[9],樊永军等对在青海云杉外生菌根中发现Tomentella 菌根真菌[10],王琴等对油松上的5 种Tomentella 外生菌根进行描述[11],魏杰等在兴安落叶松、白桦、沙地云杉、山杨和樟子松等内蒙古地区的主要树种上都发现有Tomentella 外生菌根[12-13],但对棉革菌形成的子实体鲜有报道。虽对棉革菌子实体的报道较少,但其分布极其广泛,几乎遍布全球[14-20]。
雷公山国家级自然保护区位于贵州省黔东南州中部,属于中亚热带季风山地湿润区,为贵州省苗岭山脉东段主峰,雷公山最高峰海拔为2 178.8 m,跨 度为108°5′~108°24′E,26°15′~ 26°32′N,总面积 47 792 hm2,年平均气温为14.3 ℃,年降水量 1 300~1 600 mm,具有夏无酷暑、冬无严寒、雨量充沛的气候特点,光、热、水资源丰富,气候类型多样,蕴藏着丰富的生物资源,其中高等植物278 科2 582 种,列为国家重点保护的有红豆杉、南方红豆杉、异形玉叶金花,伯乐树、秃杉、十齿花等32 种。雷公山已知的大型真菌有50 科263 种其中白蘑科、红菇科、侧耳科和多孔菌科的种类及数量占优势,占已知总数的47%,但未见棉革菌子实体的报道。
本研究从雷公山自然保护区发现的棉革菌子实体入手,通过对其进行分类鉴定,并结合分子鉴定,首次对雷公山国家级自然保护区的棉革菌子实体进行报道。
1 材料与方法
1.1 供试材料
于2016年 6—8月在雷公山国家级自然保护区的不同林分中采集样品,记录所采集到新鲜子实体的宏观形态,在常绿阔叶林(以栲树Castanopsis fargesii Franch.、青冈Quercus glauca Thunb.为优势树种)中采集子实体样本6 份,在落叶阔叶林(以亮叶桦Betula luminifera H.Winkl.、响叶扬Populus Adenopoda Maxim.为优势树种)中采集子实体样本11 份,在针叶林(以马尾松Pinus massoniana Lamb 为优势树种)中采集子实体样本9 份,在灌木从(以白栎Quercus fabri Hance 及麻栎Quercus acutissima Carruth.为优势种)中采集子实体样本3份,共计采集得子实体样本29 份。
1.2 实验方法
1.2.1 子实体样品的处理
将样品带回实验室及早加热干燥后存于锡箔纸制成的密封袋中并编号。干燥后的标本将用于解剖描述和分子鉴定。
1.2.2 子实体微观形态观察
在载玻片上分别滴2.5%的KOH 和蒸馏水,将子实体要观察的部位于解剖镜下刮下,放置于载玻片上的溶液中,盖上盖玻片,采用光学显微镜观察,对子实层中根状菌索、Subicular 菌丝、Subhymenial 菌丝、囊状体、担子及担孢子,的特征观察、照相,再进行描述及测量。子实体担孢子以及根状菌索等结构的形态使用扫描电子显微镜观察、照相,再进行描述及测量。
1.2.3 子实体样品的分子鉴定
1.2.3.1 DNA 的提取
子实体样品基因组DNA 的提取,采用试剂盒提取,采用的试剂盒为TaKaRa MiniBEST 植物基因组DNA 提取试剂盒。
1.2.3.2 PCR 扩增
PCR 反应体系为25 μL,其中加入DNA 模板2 μL,引物ITS1F(CTTGGTCATTTAGAGGAAGTAA) 0.5 μL,引 物ITS4(TCCTCCGCTTATTGATATGC) 0.5 μL,dNTP-Mix2 μL,Easy Taq buffer 2.5 µL,Taq-Polymerase 0.25 µL,ddH2O 17.25 µL。扩增程序为:94 ℃预变性5 min,94 ℃变性30 s,54 ℃退火 30 s,72 ℃延伸 45 s,35 个循环,72 ℃平展5 min。
1.2.3.3 测序
将扩增所得产物测序,测序由金斯瑞生物科技公司完成。测序后所得的序列采用Bioedit 编辑,在NCBI 中进行BLAST 序列比对。
1.2.4 系统发育分析
运用贝叶斯法对子实体样品中所获得的ITS 区段的碱基序列进行系统发育分析,构建系统发育树。
2 结果与分析
2.1 不同类型子实体形态特征描述
通过对子实体的宏观及微观形态特征进行观察分析,将初步确定为棉革菌的子实体样本分为7种类型,分别描述如下:
1)T.sp1 形态特征描述
T.sp1 子实体呈平伏状,紧贴于水青冈的腐木表面生长。子实层颜色为棕色,子实层表面为颗粒状,菌丝层颜色为深棕色。根状菌索较大,颜色为浅棕色,单型(图1A、F),菌丝紧密缠绕而且具锁状联合,很少出现单隔,直径为3.0~4.3 μm。Subicular 菌丝稍弯曲(图1B),末端具有螺纹结构(图1G),颜色为棕色,具锁状联合,无被壳,直径为3.3~4.7 μm。Subhymenial 菌丝弯曲,无短小膨大(图1C),颜色为浅棕色,具锁状联合,直径3.5~5.8 μm。无囊状体。担子形状为棒状,具有4 个小梗,不具柄,稍弯曲,颜色为浅棕色,基部具有锁状联合,无横隔,无油滴(图1D、H)。长20~28 μm。孢子形状正面观球形,侧面观椭球形,具较密集刺状纹饰,颜色为浅棕色。无油滴(图1E、I),正面观直径5.1~8.3 μm,侧面观直径5.1~8.2 μm,无厚垣孢子。
T.sp1 子实体样本采集于常绿阔叶林中。
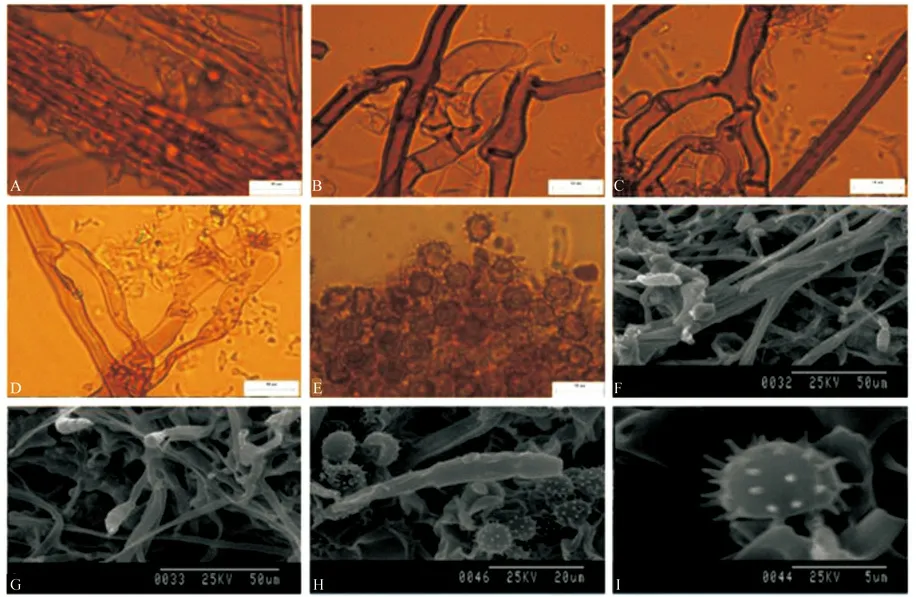
图1 T.sp1 宏观及微观形态解剖结构图Fig.1 Macroscopic and microcosmic anatomical characteristics of T.sp1
2)T.sp2 形态特征描述
T.sp2 子实体呈平伏状,紧贴于亮叶桦的腐木表面生长。子实层颜色为黑棕色,子实层表面为颗粒状,菌丝层颜色为深棕色,无根状菌索。Subicular 菌丝有短小的侧枝(图2A),颜色为深棕色,具锁状联合,少有单隔,无被壳,浅棕色,直径为4.5~7.1 μm。Subhymenial 菌丝无短小膨大(图2B),颜色为深棕色,具锁状联合,无被壳,直径4.7~6.7 μm。无囊状体。担子形状较弯曲,为瓶状,颜色为深棕色,具有4 个小梗,不具柄,担子的基部具锁状联合,偶尔具横隔(图2C、D),长24~36 μm 孢子正面观球形,倒三角形,侧面观椭球形,具有多分叉而且分离的刺状纹饰,无油滴(图2E、F),正面观直径6.3~8.8 μm,侧面观直径6.4~8.1 μm,颜色为深棕色。无厚垣孢子。
T.sp2 子实体样本采集于落叶阔叶林中。
3)T.sp3 形态特征描述
T.sp3 子实体呈平伏状,紧贴于亮叶桦的腐木表面生长。子实层颜色为黄棕色,子实体表面为绒毛状,菌丝层颜色为黄棕色。根状菌索颜色为深棕色,具有单型,二型两种,单型根状菌索具锁状联合,少具单隔,直径1.7~3.1 μm。二型根状菌索呈平行状紧密缠绕较细的骨架菌丝,不具锁状联合(图3A)。Subicular 菌丝颜色为浅棕色,具锁状联合,无被壳(图3B),直径为4.5~6.1 μm。Subhymenial 菌丝颜色为浅棕色,具锁状联合,无被壳,无短小膨大(图3C),直径4.5~6.3 μm。无囊状体。担子形状为棒状,具有4 个小梗,小梗较长,具柄,稍弯曲,基部具锁状联合,无横隔,无油滴(图3D),颜色为深黄色,长34~47 μm。孢子较大,正面观,侧面观均为球形或椭球形,颜色为棕色(图3E),具有较密集的刺状纹饰(图3F),正面观直径8.8~13.6 μm,侧面观直径8.3~10.0 μm。无厚垣孢子。
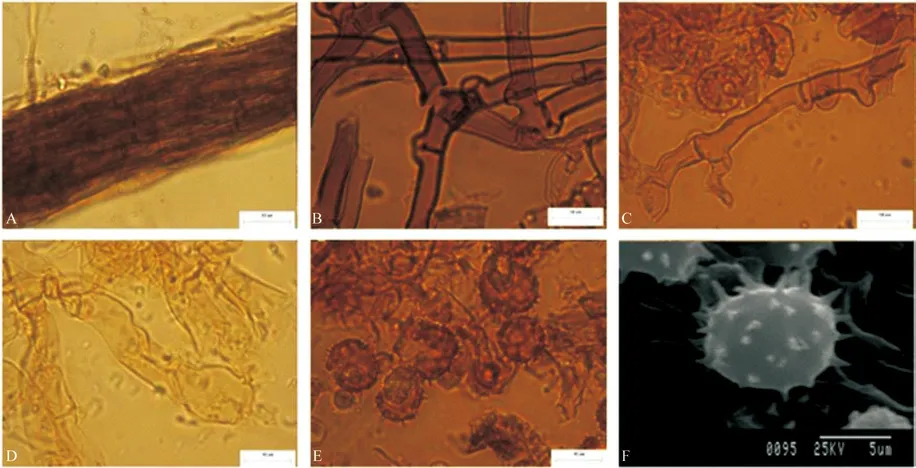
图3 T.sp3 宏观及微观形态解剖结构图Fig.3 Macroscopic and microcosmic anatomical characteristics of T.sp3
T.sp3 子实体样本采集于落叶阔叶林中。
4)T.sp4 形态特征描述
T.sp4 子实体呈平伏状,紧贴于响叶杨的腐木表面生长,子实层颜色为棕色,子实层表面为颗粒状,菌丝层颜色为棕色。根状菌索类型为单型,颜色为深棕色,具锁状联合(图4A),菌丝紧密缠绕。Subicular 菌丝有时具有被壳,颜色为棕色,具锁状联合(图4B、C)直径为3.8~5.4 μm。无囊状体。担子形状较弯曲,为棒状颜色为棕色,具柄,具有4 个小梗,担子的基部具有锁状联合(图4D),长27~33 μm。孢子正面观,侧面观均为球形至椭球形,具有密集且较短刺状纹饰,颜色为浅棕色,无油滴,(图4E、F),正面观直径5.5~7.5 μm,侧面观直径5.5~7.3 μm。无厚垣孢子。
T.sp4 该子实体样本采集于落叶阔叶林中。

图4 T.sp4 宏观及微观形态解剖结构图Fig.4 Macroscopic and microcosmic anatomical characteristics of T.sp4
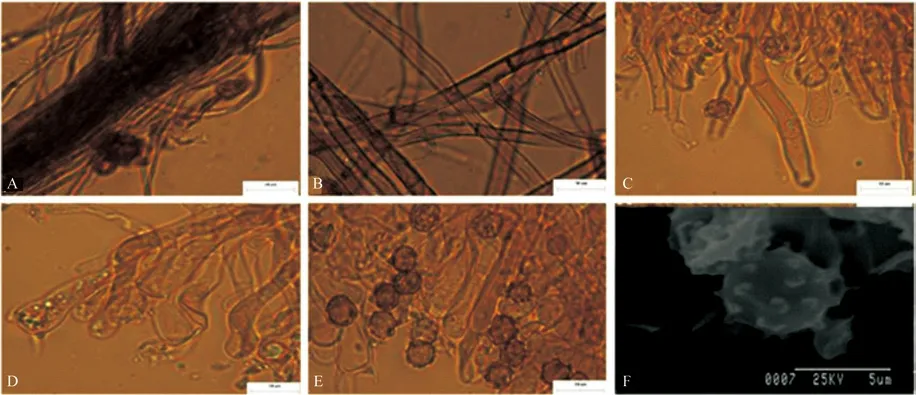
图5 T.sp5 宏观及微观形态解剖结构图Fig.5 Macroscopic and microcosmic anatomical characteristics of T.sp5
5)T.sp5 形态特征描述
T.sp5 子实体呈平伏状,紧贴于马尾松的腐木表面生长,子实体表面有蛛网膜,子实层颜色为黄棕色,子实层表面为颗粒状,菌丝层颜色为黑褐色。根状菌索类型为单型,具单隔,缠绕较松散,颜色为深棕色(图5A),菌丝直径为3.1~4.2 μm。Subicular 菌丝有时具被壳,具单隔,偶有锁状联合(图5B),颜色为棕色,直径3.5~5.4 μm。囊状体在菌丝中出现,形状为棒状,偶有被壳,具横隔(图5C)颜色为浅棕色,长47.1~83.8 μm。担子形状为棒状,不具柄,具有四个小梗,较弯曲,具锁状联合,偶有横隔出现,具油滴,常无色(图5D),长2 435 μm。孢子正面观球形,侧面观椭球形,具密集、较短、多分叉刺状纹饰,无油滴,颜色为深黄色(图5E、F),正面观直径5.0~7.1 μm,侧面直径5.3~6.7 μm,无厚垣孢子。
T.sp5 子实体样本采集于针叶林中。
6)T.sp6 形态特征描述
T.sp6 子实体呈平伏状,紧贴于马尾松的腐木表面生长。子实层颜色为深棕色,子实层表面为颗粒状,菌丝层颜色为深棕色。无根状菌索。Subicular 菌丝具被壳,少见单隔,具锁状联合,颜色为棕色(图6A、E),直径为5.0~7.2 µm。Subhymenial菌丝具被壳。无短小膨大,具锁状联合,颜色为深棕色(图6B),直径为5.2~7.4 µm。无囊状体。担子长形状为瓶状,不具柄,有四个小梗,较弯曲,具锁状联合,偶有横隔,无油滴,颜色为棕色(图6C),长为28~32 µm。孢子正面观,侧面观均为球形至椭球形,具密集的刺状纹饰,无油滴,颜色为棕色(图6D、F),正面观直径6.3~8.1 µm,侧面直径6.1~7.7 µm,无厚垣孢子。
T.sp6 子实体样本采集于针叶林中。
7)T.sp7 形态特征描述
T.sp7 子实体呈平伏状紧贴于麻栎灌木层根部的腐木上生长,子实层颜色为棕色,子实层表面为颗粒状,菌丝层颜色为棕色。根状菌索较大,单型,具被壳,具锁状联合,菌丝松散缠绕,颜色为深棕色(图7A),中央菌丝较粗,直径为4.3~8.5 µm,外围菌丝较细,1.1~2.8 µm。Subicular 菌丝具被壳,具锁状联合,(图7B、F),颜色为深棕色,直径为5.0~7.3 µm。Subhymenial菌丝偶有被壳,具锁状联合,偶有短小,但无膨大(图7C)。颜色为浅棕色,直径4.1~6.3 µm。无囊状体。孢子正面观,侧面观均为球形至椭球形,具较密集刺状纹饰,无油滴,颜色为棕色直径6.5~8.2 µm,侧面直径7.1~8.3 µm,(图7E),无厚垣孢子。
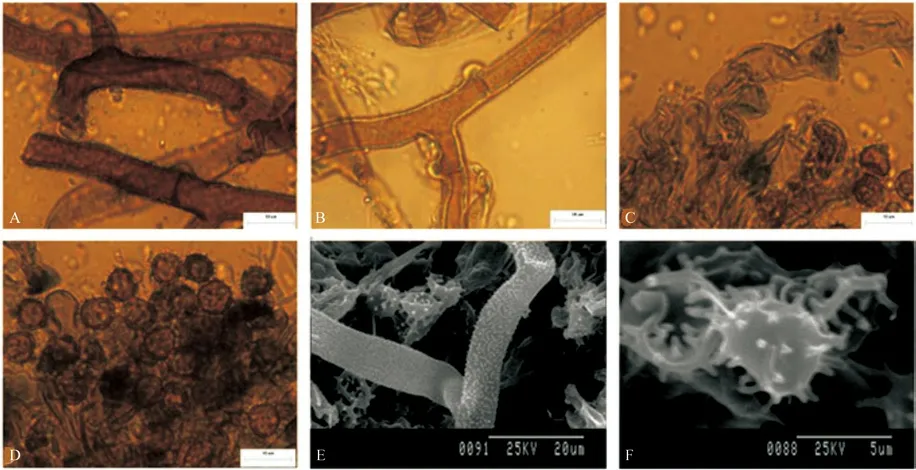
图6 T.sp6 宏观及微观形态解剖结构图Fig.6 Macroscopic and microcosmic anatomical characteristics of T.sp6
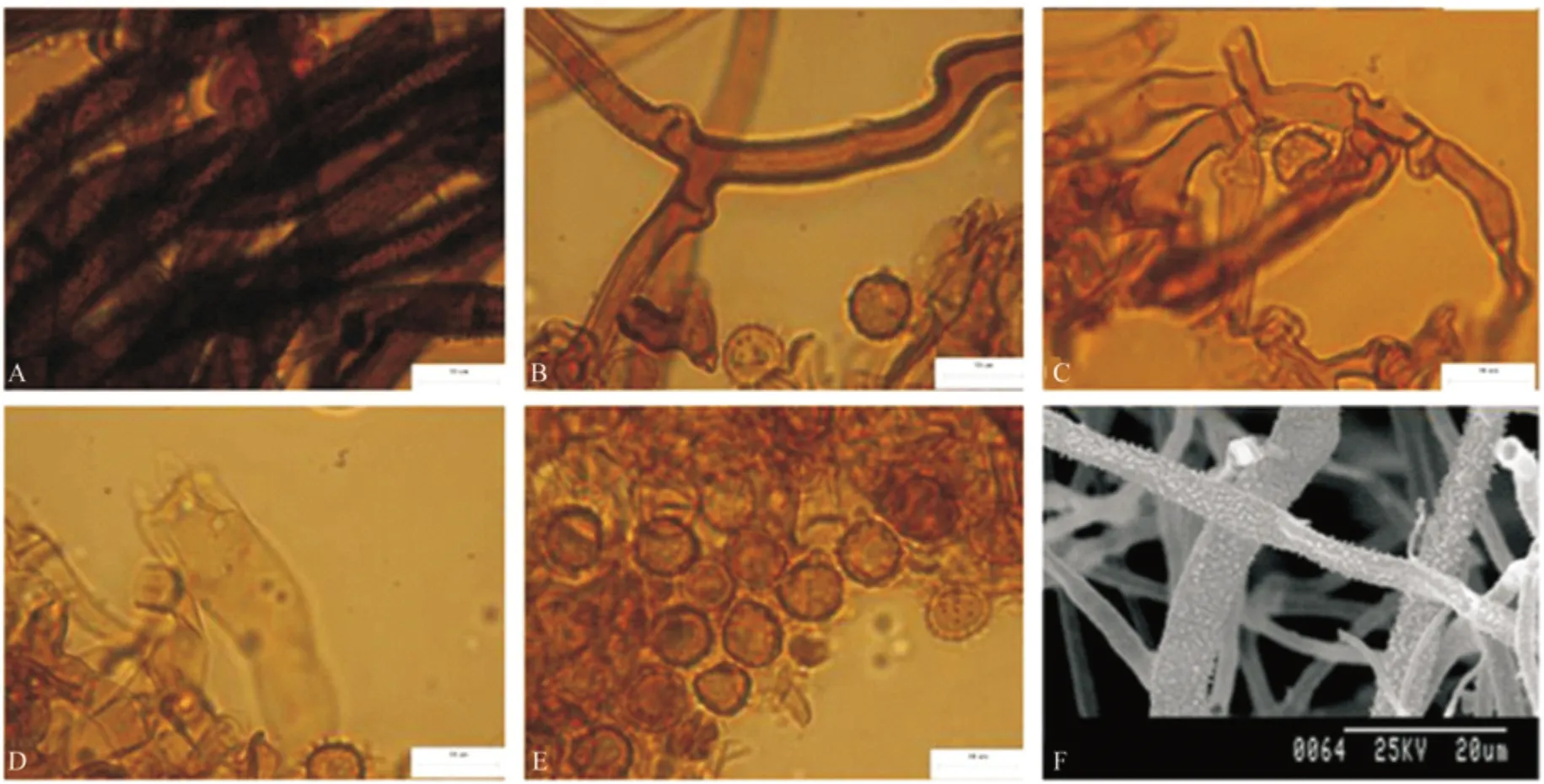
图7 T.sp7 宏观及微观形态解剖结构图Fig.7 Macroscopic and microcosmic anatomical characteristics of T.sp7
T.sp7 子实体样本采集灌木从中。
综合上述子实体宏观及微观形态特征分析可知:上述所采集的7 种类型的子实体可初步判定属于棉革菌。对其总结得出:雷公山国家级自然保护区所采集的7 种类型的棉革菌子实体的子实层颜色与菌丝层颜色基本一致,均接近棕色,7 种类型的子实层表面形状有6 种为颗粒状,只有1种为绒毛状。7 种类型的棉革菌子实体中的5 种类型具有根状菌索,根状菌索大部分为单型,其中一种类型的根状菌索具有单型、二型两种,均具锁状联合或具单隔,表面有菌丝缠绕。7 种类型的棉革菌子实体的Subicular 菌丝,Subhymenial 菌丝均具单隔或具锁状联合,较弯曲,较短小,颜色均靠近棕色。仅有一种类型出现囊状体。7 种类型的棉革菌子实体的担子颜色均与棕色相似,形状以棒状为主少数为瓶状,均具锁状联合或单隔,具柄,具横隔,较弯曲。7 种类型的棉革菌子实体的孢子形状多为球形或椭球形,纹饰类型为规则刺状。
2.2 系统发育分析
对雷公山国家级自然保护区所采集的样品进行形态学分类后获得的7 种类型的子实体分别用试剂盒提取DNA,对所获得的DNA进行PCR扩增,扩增产物再进行测序,对所得序列进行BLAST 比对,对与其同源性较高的代表序列进行下载,以Th.pseudoterrestris 为外群,将所测序列编号并与下载序列一并建立贝叶斯系统发育树(图8)。由图8分析可知,7 种类型子实体样品的序列均聚在了棉革菌Tomentella 属真菌的序列中,由此可知:此7 种类型子实体都属于棉革菌属。T.sp7 形成一个独立分支,没有与其他棉革菌序列聚集,由此推测:T.sp7 可能为尚未报道的棉革菌新种。T.sp6聚于T.ramosissima(JX630859,来自美国)与T.lapida(KU525697,来自波兰)形成的分支之间。T.sp5 与T.fuscocinerea(GU214810,来自爱沙尼亚)聚于一个分支上。T.sp4 和T.intsiae(NR121286,来自美国)聚在一起。T.sp3 与T.bryophila 相聚在一起。T.sp2 与T.stuposa(JQ888213,来自拉脱维亚)聚在一起。T.sp1 与T.galzinii(AM924141,来自俄罗斯)聚在一个分支上。说明其亲缘关系可能较近。通过采用表型分析和rDNA-ITS 序列分析等首次对雷公山国家级自然保护区7 种类型的棉革菌子实体的形态特征进行详细描述并构建系统发育树,分析得出该7种类型的子实体均属于棉革菌属。本研究采用传统分类方法,依据子实体的形态特征将雷公山国家级自然保护区采集子实体大致分为7 种不同的类型,再对其进行分子鉴定,并进行系统发育分析。通过传统的形态解剖特征鉴定以及系统发育分析研究相结合对雷公山国家级自然保护区所采集的棉革菌子实体进行研究后得知:系统发育树中部分子实体序列与数据库中的棉革菌序列聚集在一起,但对于聚集在一起的两种类型的形态解剖特征分析却发现二者存在较大差异,难以一一对应。如在系统发育树上与T.sp1 最为相近的是T.galizinii[15],但通过形态解剖特征对比发现,T.galizinii 的最为典型的特征是末端具有形状较尖的囊状体,而T.sp1 无囊状体;T.galizinii 无根状菌索,而T.sp1 具有较大的根状菌索;T.galizinii 的孢子多为掌状或倒三角形状,而T.sp1 的孢子多为球形或椭球形。出现此类现象,其原因有以下两点:第一,本次研究所采集的棉革菌子实体均是呈平伏状附着于腐木上,一般情况下难以发现,所采集的部分样品出现缺失的状况,导致对子实体标本的研究内容不够全面,程度不够深入;第二,基于数据库中所收集的绝大多数棉革菌的基因序列均来自欧美地区,缺乏来自中国地区的棉革菌序列,碱基差异通常会比较大,以往对棉革菌的研究中,由于实验条件的局限性,诸多研究均只对棉革菌属的大型真菌的形态特征进行描述,并无分子鉴定,造成基因数据库中相关棉革菌序列的缺乏,从而导致在后续的研究过程中进行序列比对时造成缺失,对系统发育分析造成了一定的困难,很多的子实体样本只能定到属的水平而不能定到种的水平。后续研究过程中应对棉革菌子实体的样品采集范围扩大,所得序列均上传至数据库中,多补充中国范围内棉革菌序列,以期推动我国对棉革菌更深层次的研究。
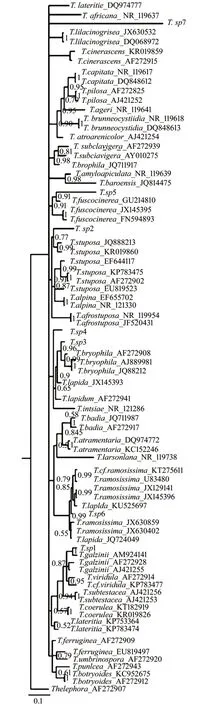
图8 基于rDNA-ITS 序列采用贝叶斯法构建所得的系统发育树Fig.8 Phylogenetic tree of samples generated from rDNAITSITS sequences by using MrBayes
