黄淮麦区34个小麦主栽品种(系)抗条锈病基因推导
2019-06-11黄亮刘太国刘博高利
黄亮 刘太国 刘博 高利

摘要为明确黄淮麦区小麦主栽品种抗条锈病基因组成,选用来自国内外的15个条锈菌菌株和21个以Avocet*S为遗传背景的小麦抗条锈病近等基因系,对我国黄淮麦区34个小麦主栽品种(系)进行苗期抗条锈病基因推导,并结合系谱分析明确其抗条锈病基因携带情况。结果表明,Yr3、Yr4、Yr8、Yr9、Yr17、Yr26、Yr27、Yr30、YrA、YrSp、YrSk分别以单基因或基因组合的形式存在于19个小麦品种中,Yr9比例最高,占29.4%,‘泰农18等9份品种仅含一个抗条锈病基因,‘山农483等10份品种含多个已知或未知基因,其余品种含有未知基因。该结果将为黄淮麦区小麦品种的合理布局提供依据。
关键词小麦条锈病;基因推导;抗条锈病基因
中图分类号:S 435.121.42
文献标识码:A
DOI:10.16688/j.zwbh.2018070
小麦条锈病是由禾谷丙锈菌小麦专化型Puccinia striiformis f.sp. tritici(Pst)引起的一种世界性小麦真菌病害,病害流行年份导致小麦严重减产乃至绝收。我国是小麦条锈病发生面积最大、危害损失最重的产麦国家[1],新中国成立以来先后发生了9次小麦条锈病大流行,其中有4次分别造成小麦产量损失600万、320万、180万和130万t[2]。条锈菌毒性的不断变异和单个抗性基因的过度使用是造成小麦条锈病大暴发的主要原因[3],选育并合理运用多基因聚合的广谱持久抗病良种是防治小麦条锈病最经济、环保和有效的方法[4]。
基因对基因学说的提出为小麦抗病基因遗传研究奠定了良好基础[5],Dubin等[6]总结了6条关于小麦抗条锈病基因推导的基本原则,为该理论在小麦抗条锈病基因推导领域的应用提供了重要指导。基因推导克服了传统方法配置杂交组合和遗传分析等费时费力的弊端,一般在4~8周就能得到有用的遗传信息,是摸清品种的抗病基因组成的有效手段。我国众多学者先后运用已知基因载体品种与条锈菌的互作关系推导出了部分小麦品种的抗条锈病主效基因,为抗条锈病基因的合理布局提供了理论依据。张玉薇等[7]推导了2006-2010年国家审定的75个小麦品种的抗条锈病基因,认为多数品种含有已知或未知抗条锈病基因,其中含Yr1、Yr2、Yr3、Yr9等基因的品种所占比例最高。王吐虹等[8]推导了我国条锈菌菌源地甘肃南部40个小麦生产品种的抗条锈病基因,发现主要以Yr9、Yr24和Yr26基因为主。
2009年在我国四川首次出现了对重要小麦抗条锈病基因Yr26和Yr10具有联合毒性的新条锈菌致病类群G22(代号V26)[9]。2016年该致病类群中流行频率最高的G229致病类型被正式命名为CYR34号生理小种[10],CYR34具有超越CYR32和CYR33成为我国第一优势小种的趋势[11],严重威胁我国小麦生产安全。黄淮麦区是我国小麦的主产区,摸清当前黄淮麦区小麦主栽品种的抗条锈病基因组成情况,及时调整小麦品种布局,预防小麦条锈病的大范围暴发成为需要解决的首要问题。基于此,我们对目前我国黄淮麦区34份小麦主栽品种进行了抗条锈病基因推导,旨在为这些品种合理布局提供科学依据。
1材料与方法
1.1材料
供试小麦为我国黄淮麦区34个小麦主栽品种(表1),由山东农业大学作物生物学国家重点实验室提供;21个以Avocet*S为背景的小麦抗条锈近等基因系,即‘Avocet*S、‘Avocet*S/Yr1、‘Avocet*S/Yr3、‘Avocet*S/Yr4、‘Avocet*S/Yr5、‘Avocet*S/Yr6、‘Avocet*S/Yr7、‘Avocet*S/Yr8、‘Avocet*S/Yr9、‘Avocet*S/Yr10、‘Avocet*S/Yr15、‘Avocet*S/Yr17、‘Avocet*S/Yr24、‘Avocet*S/Yr25、‘Avocet*S/Yr26、‘Avocet*S/Yr27、‘Avocet*S/Yr30、‘Avocet*S/Yr32、‘Avocet*S/YrA、‘Avocet*S/YrSp、Avocet*S/YrSk由澳大利亚悉尼大学植物育种研究所培育,中国农业科学院植物保护研究所收集、繁殖、保存;对照品种‘铭贤169由中国农业科学院植物保护研究所收集、繁殖、保存。
供试的15个条锈菌生理小种中,来自国外的8个,即66E74(DK)、98E26(DK)、66E10(DK)、66E154(DK)、86E90(UK)、70E72(DK)、114E26(DK)、98E90(DK);来自国内的7个,即66E26、72E66、70E10、111E239(CYR32)、111E222(CYR33)、127E255(CYR34)、63E254(G2214),所有菌种均由中国农业科学院植物保护研究所麦类真菌病害研究室保存。
1.2方法
將所有小麦品系成套播种于20 cm×30 cm的塑料盒中,用30孔的模板打穴,按照编号将近等基因系和待测小麦品种催芽后依次播种于穴内,每穴6~8粒,覆土、浇水、盖膜,置于育苗间内培养,2~3 d 后待所有小麦发芽后揭去薄膜,继续培养3~4 d,待小麦第一片叶完全展开时分生理小种进行人工接菌。接种采用刘太国等[12]的方法:每10 mg 条锈菌新鲜夏孢子加1 mL 矿物油(Soltrol170)配制孢子悬浮液,随后均匀喷洒在小麦叶片上,4~6 h后待矿物油完全晾干,将0.001%吐温水溶液均匀喷洒在叶片表面,放入洗净的接种桶中盖上薄膜,置于保湿间(黑暗、10℃),保湿24 h 后取出,人工温室(14~18℃,光照16 h/d)培养15 d,待对照品种‘铭贤169 和‘Avocet*S完全发病后调查。按照0~9 级[13]标准进行调查,即0~6 为低侵染型,7~9 为高侵染型。根据基因对基因学说和Dubin等[6]提出的6条基因推导原则,比较所有参试条锈菌生理小种对待测品种和近等基因系的侵染型,并参考系谱分析待测品种携带的抗条锈病基因。
2结果与分析
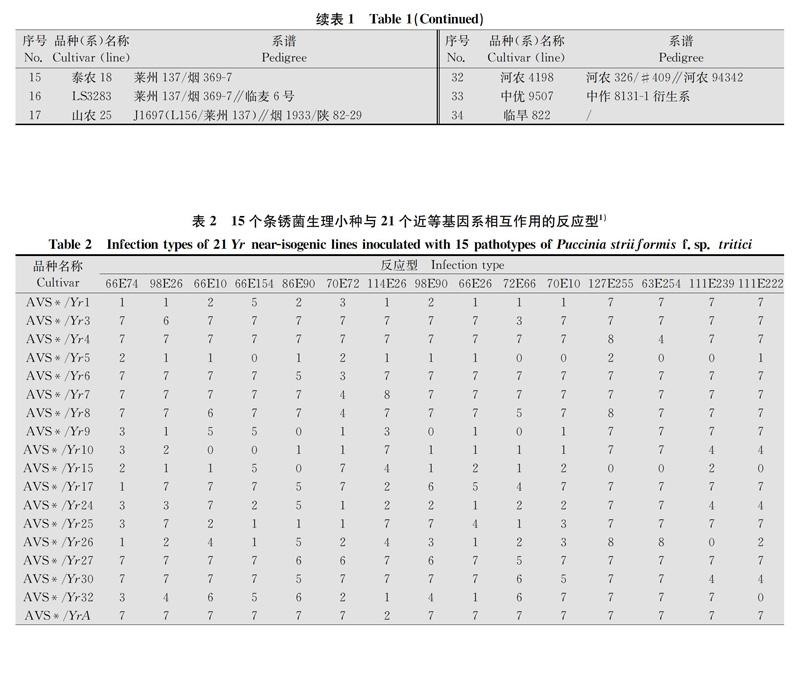
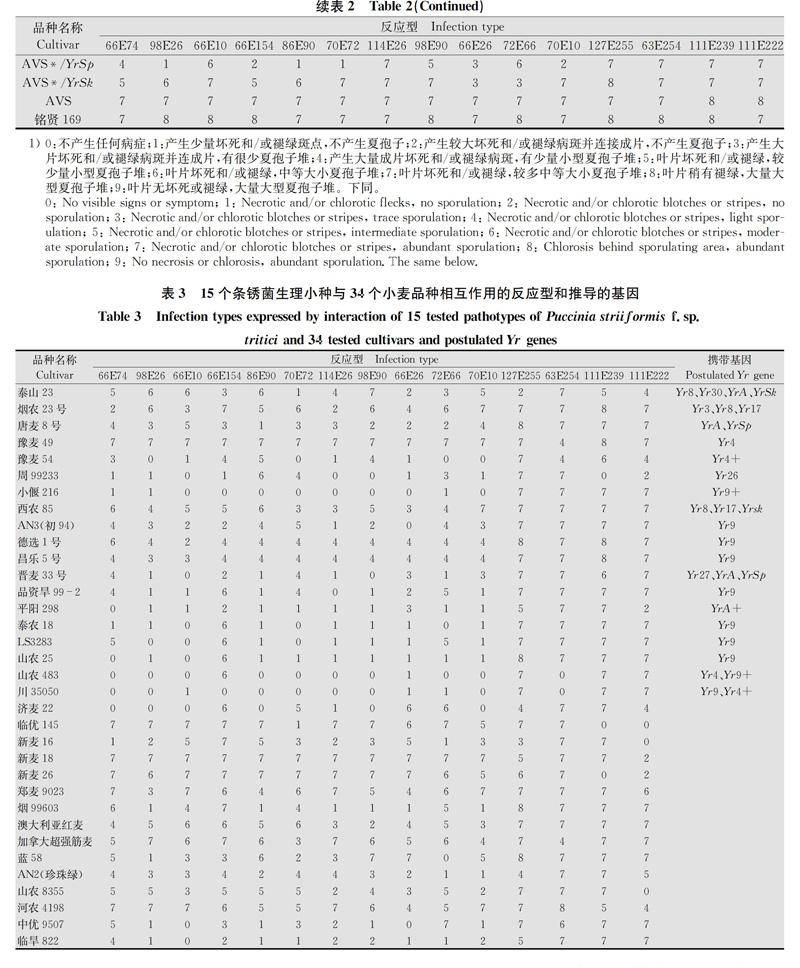
由表2、表3结果看出, ‘初94、‘德选1号、‘昌乐5号、‘品资旱992、‘泰农18、‘ LS3283、‘山农25等与Yr9抗谱一致,推测这7个品种含有Yr9基因。‘小偃216含有Yr9和其他抗性基因。Yr4仅对菌株63E254(G2214)具有抗性,‘豫麦49含有Yr4,‘豫麦54含有Yr4和其他抗性基因。 ‘山农483和‘川35050含有Yr9、Yr4和其他抗性基因。 ‘唐麦8号可能含有YrA和YrSp。‘烟农23可能含有Yr3、Yr8和Yr17基因,‘泰山23可能含有Yr8、Yr30、YrA、YrSk,‘西农85可能含有Yr8、Yr17、YrSk,‘晋麦33可能含有Yr27、YrA、YrSp,‘周99233可能含有Yr26,‘平阳298可能含有YrA和其他抗性基因。另外,‘济麦22、‘临优145、‘新麦16、‘新麦18、‘新麦26、‘郑麦9023、‘烟99603、‘澳大利亚红麦、‘加拿大超强筋麦、‘蓝58、‘AN2(珍珠绿)、‘山农8355、‘河农4198、‘中优9507、‘临旱822等15个品种与供试的已知基因的抗谱均不一致,推测这些品种可能含有供试基因之外的其他抗性基因。
1) 0:不产生任何病症;1:产生少量坏死和/或褪绿斑点,不产生夏孢子;2:产生较大坏死和/或褪绿病斑并连接成片,不产生夏孢子;3:产生大片坏死和/或褪绿病斑并连成片,有很少夏孢子堆;4:产生大量成片坏死和/或褪绿病斑,有少量小型夏孢子堆;5:叶片坏死和/或褪绿,较少量小型夏孢子堆;6:叶片坏死和/或褪绿,中等大小夏孢子堆;7:叶片坏死和/或褪绿,较多中等大小夏孢子堆;8:叶片稍有褪绿,大量大型夏孢子堆;9:叶片无坏死或褪绿,大量大型夏孢子堆。下同。
0: No visible signs or symptom; 1: Necrotic and/or chlorotic flecks, no sporulation; 2: Necrotic and/or chlorotic blotches or stripes, no sporulation; 3: Necrotic and/or chlorotic blotches or stripes, trace sporulation; 4: Necrotic and/or chlorotic blotches or stripes, light sporulation; 5: Necrotic and/or chlorotic blotches or stripes, intermediate sporulation; 6: Necrotic and/or chlorotic blotches or stripes, moderate sporulation; 7: Necrotic and/or chlorotic blotches or stripes, abundant sporulation; 8: Chlorosis behind sporulating area, abundant sporulation; 9: No necrosis or chlorosis, abundant sporulation.The same below.
3讨论
本研究结果显示,从参试的34份小麦品种中共推导出11个基因,这些基因以单基因或基因组合的形式存在于19个小麦品种中,部分品种间抗病谱存在细微差异,其中10个品种携带Yr9,3个品种携带Yr8,携带Yr4和YrA的品种各4份,携带Yr17、YrSp和YrSk的品种各2份,携带Yr3、Yr26、Yr27和Yr30基因的品种各1份。其中仅含1个基因的品种共9份,含一个及以上已知或未知基因的共10份,有15个品种未能推导出含有已知基因但对部分小种依然具有较高的抗性,其抗性可能是由其他抗性基因决定的。
Yr9对CYR32、CYR33、CYR34、G2214表现感病,对其余的参试菌株表现抗病,在所有参试的34份小麦中Yr9基因所占的比例最高,达到了29.4%,其原因是Yr9基因位于小麦1BL/1RS易位系上,该易位系具有抗病性、丰产性和稳产性,于20世纪后期引入我国并在生产中大量使用[14],所以我国目前的小麦品种大多含有该易位系。黄亮等[15]、李敏州等[16]的研究结果中Yr9基因的含量均超过30%,与本研究结果基本保持一致。根据系谱分析发现,‘泰农18(莱州137/烟3697)、‘ LS3283(莱州137/烟3697∥临麦6号)、‘山农25[J1697(L156/莱州137)∥烟1933/陕8229]3个品种均具有‘莱州137的血统,且‘莱州137为‘洛夫林10和‘洛夫林13的衍生品种,带有小麦1BL/1RS易位系[15],所以推测它们的Yr9基因来自于‘莱州137;‘山农483为‘矮孟牛的衍生品种,而‘矮孟牛为亲本‘牛朱特的子1代,所以推测‘山农483的Yr9基因來源于‘牛朱特;‘小偃216[(郑引4号/小偃96×7751)/小偃107∥兰考906]具有 ‘兰考906的遗传背景,‘兰考906又名‘豫麦66,是通过小麦细胞工程选育的具有六倍体小黑麦血统的高产品系,所以推测‘小偃216的Yr9基因源自于‘兰考906。
黄淮麦区是我国的主要产麦区,小麦的种植面积和产量分别占全国的45%和51%[17],享有我国“粮仓”的美誉,小麦条锈病是该地区小麦生产的主要病害之一,每年都有不同程度的发生,影响小麦产量和品质。本试验参试品种‘郑麦9023({(小偃6号/西农65)∥[83(2)33/84(14)43]}F3/陕213)和‘济麦22(临远7069/鲁麦14∥935106)是黄淮麦区重要的主推品种,每年的种植面积均领先其他小麦品种,虽然本试验在这两个品种中并没有推导出已知抗性基因,但是研究发现这两个品种对部分参试菌株具有较好的抗性,初步认为这两个品种可能含有其他未检测到的抗条锈病基因,‘济麦22具有‘鲁麦14的血统,研究表明‘鲁麦14含有未知抗条锈基因[18],所以‘济麦22的未知抗条锈基因可能来自‘鲁麦14。
植物的抗病性并不只是单基因遗传控制,更多的是基因间相互作用的结果,基因间的相互作用会造成抗病性互补、累加等优势[19]。基因间相互作用现象在本试验中也有体现,例如本试验的供试品种 ‘山农483,聚合了抗条锈病基因Yr9和Yr4之后兼具了两个基因的抗性,使‘山农483的抗性增强、抗性谱扩宽,这样的现象在其他学者的研究中也有体现,如聚合了抗条锈基因Yr1、Yr2、Yr9、Yr32的持久抗性品种‘Savannah抗性得到了明显的提高[20]。所以在育种过程中将多个抗条锈基因聚合到同一小麦品种中是增强抗病性、拓宽抗病谱的有效途径。
需要引起重视的是,参试的34份小麦品种对我国当前流行的条锈菌生理小种CYR32、CYR33、CYR34以及条锈菌致病类型G2214大多表现感病,这意味着我国黄淮麦区将持续受到小麦条锈病的威胁,在品种布局时应尽量考虑周全,将条锈病造成的危害降到最低。
本研究虽然采用基因推导的方法推测了参试品种的抗条锈病基因,但是受含抗条锈近等基因系的品种数量和小麦条锈菌的致病性等的限制,部分品种依然无法推导出可能的抗性基因,并且部分参试品种未能查到相应的系谱,影响了试验结果的准确性。所以在使用基因推导方法确定抗病基因时,应尽量多地使用已知基因载体品种和毒性差异更显著的致病菌生理小种,并且使用的菌株应涵盖更多毒性谱,在条件允许的情况下可使用分子标记、等位性测验等手段进行验证,从而提高试验结果的准确度。
参考文献
[1]陈万权,康振生,马占鸿,等.中国小麦条锈病综合治理理论与实践[J].中国农业科学,2013,46(20):42544262.
[2]WAN Anmin, CHEN Xianming, HE Zhonghu. Wheat stripe rust in China [J]. Australian Journal of Agricultural Research, 2007, 58(6): 605619.
[3]李振岐.我国小麦品种抗条锈性丧失原因及其解决途径[J].中国农业科学,1980,13(3):7277.
[4]吴立人,牛永春.我国小麦条锈病持续控制的策略[J].中国农业科学,2000,33(5):4654.
[5]FLOR H H. The complementary genic systems in flax and flax rust [J]. Advances in Genetics, 1956, 8(4): 2954.
[6]DUBIN H J, JOHNSON R, STUBBS R W. Postulated genes for resistance to stripe rust in selected CIMMYT and related wheats [J]. Plant Disease, 1989, 73(6): 472475.
[7]张玉薇,刘太国,刘博,等.中国75个国审小麦品种抗条锈基因推导[J].植物保护学报,2014,41(1):4553.
[8]王吐虹,郭青云,蔺瑞明,等.中国40个小麦农家品种和甘肃南部40个生产品种抗条锈病基因推导[J].中国农业科学,2015,48(19):38343847.
[9]LIU Taiguo, PENG Yunliang, CHEN Wanquan, et al. First detection of virulence in Puccinia striiformis f.sp tritici in China to resistance genes Yr24 (=Yr26) present in wheat cultivar Chuanmai 42 [J].Plant Disease,2010,94(9):1163.
[10]刘博,刘太国,章振羽,等.中国小麦条锈菌条中34号的发现及其致病特性[J].植物病理学报,2017,47(5):18.
[11]张勃,贾秋珍,黄瑾,等.小麦条锈菌新菌系贵229和贵2214发展趋势与毒性分析[J].西北农业学报,2015,24(7):125130.
[12]刘太国,章振羽,刘博,等.小麦抗条锈病基因Yr26毒性小种的发现及其对我国小麦主栽品种苗期致病性分析[J].植物病理学报,2015,45(1):4147.
[13]LINE R F, QAYOUM A. Virulence, aggressiveness, evolution and distribution of races of Puccinia striiformis (the cause of stripe rust of wheat) in North America, 196887 [R]. USDA Technical Bulletin No. 1788. National Technical Information Service, 1992.
[14]WIESER H, KIEFFER R, LELLEY T. The influence of 1B/1R chromosome translocation on gluten protein composition and technological properties of bread wheat [J]. Journal of the Science of Food and Agriculture, 2000, 80(11): 16401647.
[15]黄亮,刘太国,肖星芷,等.中国79个小麥品种(系)抗条锈病评价及基因分子检测[J].中国农业科学,2017,50(16):31223139.
[16]李敏州,李强,巢凯翔,等.陕西省115个小麦品种(系)抗条锈病基因的分子检测[J].植物病理学报,2015,45(6):632640.
[17]李峰奇,韩德俊,魏国荣,等.黄淮麦区126个小麦品种(系)抗条锈病基因的分子检测[J].中国农业科学,2008,41(10):30603069.
[18]牛永春,乔奇,吴立人.豫鲁皖三省重要小麦品种抗条锈基因推导[J].植物病理学报,2000,30(2):122128.
[19]李博,张志毅,张德强,等.植物杂种优势遗传机理研究[J].分子植物育种,2007,5(S1):3644.
[20]ERIKSEN L, AFSHARI F, CHRISTIANSEN M J, et al. Yr32 for resistance to stripe (yellow) rust present in the wheat cultivar Carstens V [J].Theoretical & Applied Genetics,2004,108(3): 567575.
(责任编辑:杨明丽)
