干旱荒漠区植物生态位对水盐的响应
2019-05-31郄亚栋滕德雄吕光辉
郄亚栋,滕德雄,吕光辉
1 绿洲生态教育部重点实验室,乌鲁木齐 830046 2 新疆大学资源与环境科学学院,乌鲁木齐 830046 3 新疆大学干旱生态环境研究所,乌鲁木齐 830046
生态位广泛地指在一个由一系列环境轴所定义的n维空间内种群可以维持存活的能力,能够反映物种在群落中的功能地位,并且可以定量地研究种内、种间、生境三者之间的相互关系[1- 2],已成为现在生态学研究中的一个重要理论,特别是在种间联系、群落结构和功能、生物资源供需、物种进化等方面的研究中有重要作用,为阐释自然生物群落中物种共存与竞争机制提供了夯实的根基[3- 5]。
目前,全球气候变化的持续正在强烈地影响着生态系统的功能,导致生物生境的改变,并对生物的生存已构成严重的危险,极端地情况下可能会引起全球生物多样性的丧失[6- 7]。然而,不同的物种或生态系统对生境压力的脆弱性不同,具有不同的生态位特征[3,8- 10]。以往研究表明,两个物种在竞争相同环境资源的同时,如果其他所有的生态因素不变,那么两者可能是不能共存的[8,11]。种群对资源利用能力的大小体现了自身在群落生态空间内的地位,也就是生态位[1,12]。一些学者认为,普遍的物种或生态系统都具有较广泛的环境适应范围,其生态位宽度较大,分布范围狭窄的物种或生态系统,其生态位宽度较小[13- 14]。然而,一些研究发现,物种在群落生态空间的地位是可以通过自身的生态策略扭转的,认为竞争能力较弱的物种可能会在多维的生态空间下选择其中的一小部分进行繁殖和生存,以此逃避竞争的排斥作用,随着时间的推移,物种反而会在受限制的生态空间内占据主导地位,对该资源梯度下其他物种的生态空间产生作用,甚至可能会将其他物种排除在外[12,15]。贺强等[16]在黄河三角洲湿地的水盐梯度上发现,物种出现生态位分化现象,且同资源纬度下的物种间生态位重叠较大;García-Baquero等[17]、刘扬等[18]和彭舜磊等[19]对植物生态位分析均发现,土壤水分是引起生态位分化的驱动因子;Iturrate等[20]发现土壤水分和盐分共同引起了植物生态位分化,并认为是决定冻原物种共存和群落组成的重要因子;张东梅等[21]在花马池盐湖地区发现,盐分梯度上生态位重叠值与Ochiai指数相关程度均高于水分梯度。上述对植物生态位的研究均是集中在半干旱、高寒或湿润地区,对干旱荒漠区植物生态位的研究较少,并且在干旱荒漠区由于物种稀少、群落单一和系统脆弱对环境变化响应相对敏感[22-24],土壤水分和盐分更是其植物生存和群落组成的驱动因子和限制因子。然而,整合水盐因子在自然梯度下对植物生态位研究更是缺乏系统性。
艾比湖流域,生物资源较丰富,景观类型多样,兼具荒漠生态系统和湿地沼泽生态系统的双重特征,是研究干旱荒漠区植物生态位的适合对象[24]。本文在研究干旱荒漠区土壤水分、盐分和植物生态位特征的基础上,分析了不同水盐梯度下植物的群落组成和植物生态位响应土壤水盐的变化趋势,以回答以下科学问题:(1)沿水盐梯度植物群落的特征;(2)水盐梯度下群落中植物的生态位特征;(3)干旱荒漠区植物生态位对土壤水盐响应,以期为艾比湖流域植物生态位研究提供基础科学数据,为干旱荒漠区植物的利用与保护提供理论支撑。
1 材料和方法
1.1 研究区概况
艾比湖湿地国家级自然保护区位于阿拉山口东南方向,地理坐标为44°30′—45°09′N,82°36′—83°50′E,属于湿地-荒漠森林复合生态系统类型的自然保护区。保护区总面积2670.85 km2,海拔189 m。该地区属于温带大陆性干旱气候,夏季酷热,冬季严寒,干旱少雨。1月份平均气温-16℃,极端最低温-36℃;7月份平均气温27℃,极端最高温44℃,年平均降雨量100 mm左右,年平均蒸发量1600 mm以上,全年大风日数约170d,最大风速55 m/s。区内土壤类型为灰漠土、灰棕漠土和风沙土,隐域性土壤为盐(盐渍化)土、草甸土和沼泽土。区内土壤盐渍化程度较高,其主要形成以湖区为中心向四周呈环状递减格局,或受河流和气候影响呈垂直于河的带状减少分布,类型有原生盐碱化和次生盐碱化[25]。研究区主要植物种类有胡杨(Populuseuphratica)、梭梭(Haloxylonammodendron)、白刺(Nitrariatangutorum)、琵琶柴(Reaumuriasoongorica)、盐爪爪(Kalidiumfoliatum)、骆驼刺(Alhagisparsifolia)、罗布麻(Apocynumvenetum)、花花柴(Kareliniacaspica)和芦苇(Phragmitesaustralis)等。
1.2 研究方法
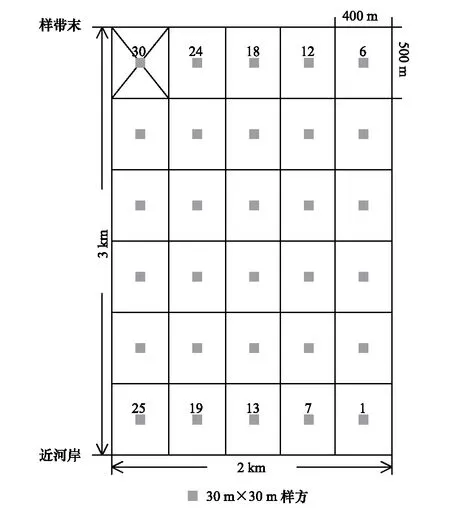
图1 样方分布图Fig.1 The location of the plots
1.2.1野外调查和样品采集
于2014年7—8月份(植物生长旺季)在艾比湖湿地保护区内东大桥管护站西北方向进行野外调查。垂直于阿其克苏河,布置2 km×3 km样地,将其等分成30个400 m×500 m小样地(图 1)。于每个小样地中心部位设置一个30 m×30 m样方(图 1),进行群落调查和样品采集。在调查样方过程中,记录每个样方中心点的地理坐标、海拔,以及样方内出现的所有物种的种类,对于不能够识别的物种,采集标本并编号,带回实验室鉴别;对于乔木,记录其多度、高度、冠幅、胸径;对于灌木和草本,记录高度、丛(株)数、丛幅、盖度。样方清查后,分别在30个样方内按照冠下土、裸地土挖取0—50 cm的土壤剖面,并将两种类型的土壤用四分法混合均匀作为该样方的土壤样品,取适量装入铝盒中,并称其重量,用于测定含水量的土壤鲜重数据;再取适量样品土壤装入密封袋内,将其带回实验室用于盐分含量的测定。
1.2.2样品分析
用烘干称重法测定土壤的含水量,用残渣烘干法(土水比1∶5)测定土壤的含盐量。
1.2.3水盐协同聚类
依据水分和盐分含量对样方进行系统聚类[26],分析水盐协同作用对群落生态位的影响。由聚类分析树的结果,将样方划分成不同群落类型,以不同群落下的样方数作为“资源状态”,计算不同物种在不同群落中的重要值,进而得出它们在不同水盐梯度下的生态位宽度以及生态位重叠,并分析它们在不同水盐梯度下群落中的功能地位以及相互之间利用资源的状态。
1.2.4物种重要值的计算
根据荒漠植物群落中植物的高度、个体数量、频度和盖度等数量指标,分别统计和计算乔木、灌木和草本物种的重要值:重要值乔木=(相对多度+相对频度+相对盖度+相对高度)/4,重要值灌木=(相对多度+相对盖度+相对频度)/3,重要值草本=(相对盖度+相对频度)/2[27]。其中,相对多度=(某种植物的多度/全部植物的多度之和)×100%,相对频度=(某种植物的频度/全部植物的频度之和)×100%,相对盖度=(某种植物的盖度/全部植物的盖度之和)×100%,相对高度=(某种植物的平均高度/全部植物的平均高度之和)×100%。
1.2.5种群生态位的研究方法
(1)生态位宽度采用Levins[28]提出后经Colwell[29]加权修改的计算方法:
式中,Bi为物种i的生态位宽度,Pij=nij/Ni,nij为物种i在资源j上的重要值,Ni表示种i在所有资源等级上的重要值之和;Pij代表物种i在资源j上的重要值占该物种在所有资源等级上的重要值总和的比例,r为样方数。
(2)生态位重叠采用Pianka生态位重叠指数[30]:
式中,Oij为物种i和j的生态位重叠指数,Pij和Pkj分别为第i和第k物种在第j个资源上的重要值,r为样方数。
1.2.6群落生态位宽度的计算方法
根据物种的重要值,对其生态位宽度测度值进行加权,把加权后的总和定义为群落的生态位宽度。
1.2.7变系数模型
利用变系数模型(Varying Coefficient Models)[31]进行群落生态位宽度对水盐响应特征的分析。模型结构如下:
Bi=α(Ui)Xi+b(Ui)+εi,i=1,2,…,n
Bj=β(Xj)Uj+b(Xj)+μj,j=1,2,…,n
式中,Bi为群落生态位宽度;Xi为土壤盐分,Ui为土壤水分,α(Ui)代表生态位盐效应指数,b(Ui)为该模型的截距项,i为样方序号;Bj为群落生态位宽度;Xj为土壤盐分,Uj为土壤水分,β(Xj)代表生态位水效应指数,b(Xj)是该模型的截距项,j为样方序号。
1.2.8数据分析
水盐聚类分析在SPSS 20.0中进行;植物生态位对土壤水盐的响应特征在R语言(Version 3.4.3,R Core Team)中采用变系数模型分析,并用ggplot2程序包进行绘图;利用Microsoft Office Excel 2013对数据进行前期整理和计算;运用Origin 8.0对物种生态位测度值作图。
2 结果和分析
2.1 植物群落调查结果及水盐聚类
样地中共有多年生木本植物9科16属16种,皆为被子植物,其中占优势科的属和种主要有豆科(3属3种)和藜科(4属4种)。为防止偶然结果,剔除偶见种3种,剩余8科13种,其中乔木1种:胡杨,小乔木2种:梭梭和柽柳(Tamarixramosissima),灌木2种:白刺和铃铛刺(Halimodendronhalodendron);半灌木5种:琵琶柴、骆驼刺、罗布麻、盐爪爪和盐节木(Halocnemumstrobilaceum);草本3种:甘草(Glycyrrhizauralensis)、芦苇和花花柴。因20和30号样方采集数据不完全而被剔除,剩余28个样方数据,基于这28个样方土壤的水分和盐分进行系统聚类,将聚类结果与实际的生态意义相结合,可划分为3个水盐梯度群丛(表1),命名结果及环境特征如下:
群丛I,胡杨-梭梭-罗布麻-芦苇(Ass.Populuseuphratica-Haloxylonammodendron-Apocynumvenetum-Phragmitesaustralis),包含样地1、2和7,主要分布于阿其克苏河近岸边及地下水较浅的地区。该植被系统的主要群落为胡杨群落,铃铛刺、琵琶柴和白刺为主要伴生种,未出现种有盐爪爪、盐节木和花花柴。该群丛土壤含水量、含盐量最高。
群丛II,盐爪爪||胡杨-梭梭-琵琶柴(Ass.Kalidiumfoliatum‖Populuseuphratica-Haloxylonammodendron-Reaumuriasoongorica),包括3—6,8—17,19,21—23和25—28共22个样方,主要分布于阿其克苏河的河滩边缘地带,13种植物均有出现,为艾比湖流域的主要荒漠植物群落,多沙土与盐渍灌丛土。植物群落以沙生植物和盐生植物为主,主要伴生种有柽柳、盐节木、花花柴和芦苇。该群丛土壤含水量、含盐量较高。
群丛III,骆驼刺||胡杨-梭梭-琵琶柴-花花柴(Ass.Alhagisparsifolia‖Populuseuphratica-Haloxylonammodendron-Reaumuriasoongorica-Kareliniacaspica),包含样地18,24和29,主要分布在距河岸带较远的荒沙土上,植物组成种类简单,植被盖度较低,柽柳和芦苇为主要伴生种,未出现种有铃铛刺和甘草。该群丛土壤含水量、含盐量最低。

表1 研究区植物群落类型及环境特征(平均值)
I:高水盐土壤环境的群落I,Community I of the high saltwater soil environments;II:中水盐土壤环境的群落II,Community II of the moderate saltwater soil environments;III:低水盐土壤环境的群落III,Community III of the low saltwater soil environments
2.2 不同水盐梯度群落下种群的生态位宽度和生态位重叠
对不同水盐梯度下的种群生态位宽度计算结果表明,I类型下,胡杨、琵琶柴和梭梭具有较高的生态位宽度值,分别为0.994、0.994和0.977,但柽柳、骆驼刺和甘草的生态位宽度值较低,均为0.333;II类型下,白刺和琵琶柴具有较高的生态位宽度,分别为0.899和0.879,而铃铛刺和甘草明显低于其他物种,分别为0.045和0.066;III类型下,梭梭、花花柴和琵琶柴具有较高的生态位宽度,分别为0.992、0.967和0.922,而盐节木的生态位宽度较低,为0.333(表2)。总的来说,不同水盐梯度下同一物种的生态位宽度存在差异,不同物种在同一水盐梯度下的生态位宽度也有所差异,其中琵琶柴在各水盐梯度均具有较高的生态位宽度值,而甘草和盐节木在各水盐梯度的生态位宽度值均较小。
对不同植物群落类型下种群生态位重叠的计算结果表明,在群落类型I中,胡杨、梭梭和琵琶柴与其他物种的生态位重叠均较高,柽柳和骆驼刺的生态位重叠值最高,为1,但它们两者与其他物种的生态位重叠值都不高,其中分别与铃铛刺和甘草的重叠值最低,均为0(表3)。在群落类型II中,白刺和琵琶柴与其他物种的生态位重叠值较高,其中白刺和琵琶柴的重叠值最高,为0.917,铃铛刺和甘草与其他物种的生态位重叠均较低,其中铃铛刺与柽柳、盐爪爪、盐节木、花花柴和甘草以及柽柳与甘草的重叠均最低,为0(表4)。群落类型III下,胡杨、梭梭、琵琶柴和花花柴与其他物种的生态位重叠值均较高,但柽柳和白刺的生态位重叠值最高,为0.998,盐节木和芦苇的重叠值最低,为0(表5)。

表2 不同水盐梯度下13种植物的生态位宽度值
符号“—”代表此水盐梯度下不存在该物种

表3 群落类型I的植物生态位重叠值
字母的含义见表2

表4 群落类型II的植物生态位重叠值

表5 群落类型III的植物生态位重叠值
综合生态位宽度和生态位重叠值的分析表明,植物群落类型I、II和III下生态位宽度大于0.8的物种分别占所有物种的60.00%,15.38%,45.45%,而低于0.2的分别占有0、23.08%、0;植物群落类型I、II和III中生态位重叠值超过0.8的种对分别占所有种的37.78%,11.54%,43.64%,而低于0.2的分别占有8.89%,24.36%,5.45%(图2),说明群落类型I和III种群的生态位宽度较大,并且具有较高的生态位重叠,而群落II种群的生态位宽度较低,其生态位重叠值也较低。在群落水平上考虑种群的生态位宽度与生态位重叠之间的关系可见(图3),两者之间在一定程度上具有显著的相关关系(R2=0.45,P<0.01),生态位宽度较大的种群,其生态位重叠值也较大,而生态位宽度较小的种群,生态位重叠值也较小。
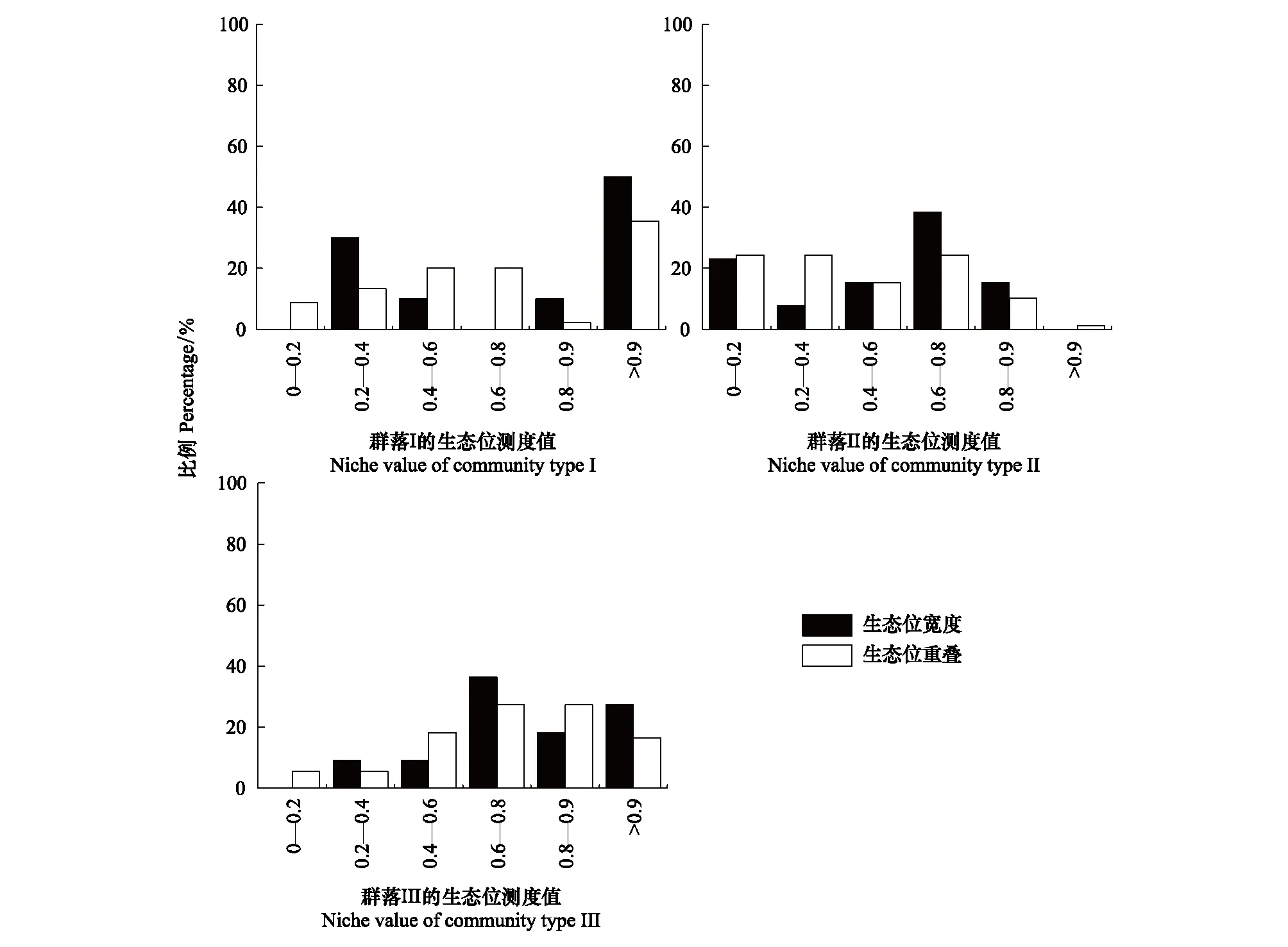
图2 研究区不同群落下物种生态位测度值的分布格局Fig.2 Distribution pattern of species niche values in different communities in the study areaI:高水盐土壤环境的群落I,Community I of the high saltwater soil environments;II:中水盐土壤环境的群落II,Community II of the moderate saltwater soil environments;III:低水盐土壤环境的群落III,Community III of the low saltwater soil environments

图3 群落水平上物种生态位测度值的相关关系 Fig.3 Correlation of species niche values at community level
2.3 群落生态位对水盐的响应特征
根据变系数模型分析群落生态位对水盐响应的结果如图4所示,其中图4,分别为模型(1)的参数项[α(Ui)]和截距项[b(Ui)]的拟合结果,分析表明,当土壤水分小于6.06%时,生态位盐效应指数[α(Ui)]小于0,说明盐分对群落生态位是呈现负效应的,反之,土壤水分大于6.06%时,生态位盐效应指数大于0,说明盐分对群落生态位是呈现正效应的,而当土壤水分等于6.06%时,生态位盐效应指数等于0(图4);如果不考虑盐分对生态位的效应时,那么群落生态位随着土壤水分的减少呈现降低的趋势(图4)。图4,分别为模型(4)的参数项[β(Xj)]和截距项[b(Xj)]的拟合结果,分析表明,当土壤盐分小于4.87 g/kg时,生态位水效应指数[β(Xj)]小于0,说明水分对群落生态位是呈现负效应的,反之,土壤盐分大于4.87 g/kg时,生态位水效应指数大于0,说明水分对群落生态位是呈现正效应的,而当土壤盐分等于4.87 g/kg时,生态位水效应指数等于0(图4);在不考虑水分对生态位效应时,群落生态位随着土壤盐分的减少呈现降低的趋势(图4)。总的来说,在高水高盐的土壤水盐分配模式下,群落的生态位受水盐的协同作用,主要表现为升高趋势;在低水低盐的土壤水盐分配模式下,群落的生态位受水盐的协同影响,主要表现为降低趋势。

图4 群落生态位对水盐的响应Fig.4 Response of community niche to water and salt
3 讨论
3.1 群丛分类及其决定因素
植物群落是植物在生存环境的长期变化中通过自身的调节机制和适应策略,而聚集在相同地段上的各物种种群的复合体形成的,是维持生态系统物质循环和能量流动的基础与动力[32]。根据艾比湖地区土壤水盐特征,将28个植被样地划分为3个主要植物群丛,其物种类型的组成(I:10种-II:13种-III:11种)沿水盐环境因子梯度呈现倒“V”型,这与Kuglerová等[33]的研究结果不一致,Kuglerová等认为,河岸带的土壤水分条件最好,其植物组成应该最为丰富;Zhu等[34]和Sara等[35]也有相似的结论,认为地下水埋深是制约群落类型、植物分布格局的主要因子,物种的丰富度与地下水位的埋深呈高度正相关。对于本研究区群落的环境特征,因艾比湖研究区处于准噶尔盆地西南缘洼地和水盐的汇聚中心,I型群丛距离阿其克苏河河岸较近,土壤获得河流的补给水分相对较多,地下水位高,而在干旱的环境下,盐分随着水分的蒸散运移至土壤表层,浅层土壤盐分集聚,形成高水高盐的水盐空间分布模式,致使植物群落面临盐碱胁迫,这对深根系植物胡杨和梭梭的生长有益,对不耐盐碱的浅根系植物不利[36],因此,河岸边呈现出以胡杨占绝对优势的胡杨群落,植物群落的物种丰富度较低;随着与河流距离的增大,土壤可获得的补给水量减少,地下水位下降,土壤表层盐分随之降低,根系较潜的盐爪爪、盐节木和花花柴等盐生小灌木和草本植物出现,其物种丰富度达到最大,在沿河梯度约1—2.5 km的区域内,土壤适宜的水分条件与较低的盐渍环境,维持着较高的物种多样性;进一步加大与河道的距离,地下水位持续下降,导致土壤含水量明显减少,不足以维持较高的物种丰富度[32-35]。
3.2 不同水盐梯度群落下种群的生态位宽度和生态位重叠
生态位宽度可以描述种群在群落中所利用资源总和的能力,其大小取决于环境因子的分配模式、物种对环境的生存力和种间的竞争强度。生态位宽度值大的物种,对环境的适应能力越强,越能充分地利用各种资源,且在群落中往往处于优势地位[2- 4]。在植物群落I类型下,土壤表层属于高水高盐的水盐空间分布模式,利于胡杨、琵琶柴和梭梭等深根系植物的生长,具有较高的生态位宽度值,但不利于柽柳、骆驼刺和甘草等不耐盐碱的浅根系植物生长,其生态位宽度值较低;植物群落II类型下,土壤水盐有所降低,一些浅根系耐盐碱植物开始生长,比如盐爪爪和盐节木,但其生态位宽度并非是最大,而白刺和琵琶柴作为研究区内盐生灌丛的建群种,分布广泛,具有一定的优势地位,生态位宽度较大,铃铛刺和甘草的生态位宽度均比较小,这说明两者分布范围比较窄,对生境的要求具有选择性;III类型植物群落下,土壤水盐含量明显减少,梭梭、花花柴和琵琶柴等旱生植物占取该群落的主要地位,其生态位宽度均较大,而盐生植物盐节木的生态位宽度较低,这说明该群落土壤的水盐环境不适合盐节木的生存。以上表明,不同的土壤水盐环境决定了物种的生态位宽度,反映了种群资源利用状况对水盐梯度的响应,表明各物种在群落中的地位,进一步明确了不同水盐梯度下植物群落的结构组成[2, 8- 9]。另一方面,本研究区种群的生态位宽度除了由不同水盐梯度下植物群落的结构组成决定以外,物种不同的生物学和生态学特性也是一个很重要的因素[12-14],如琵琶柴在3种水盐梯度下均具有较高的生态位宽度,说明其在该地区具有特殊的适应机制,叶片属于肉质短小的圆柱形,能够有效的储存水分,以及阻止水分散失,同时琵琶柴具有基生叶稠密、根系发达和簇生等特点[37],可以充分利用环境资源,在各水盐梯度下占取群落中的优势地位;而甘草以无性繁殖为主,依靠其根茎断裂脱离母体而形成,决定了对生境的要求具有一定的苛刻性,繁殖与生长受到限制,分布范围较窄,在3种水盐梯度下的生态位宽度均较低[38]。
生态位重叠反映两个或两个以上生态位相似的物种对同一空间资源利用和竞争的力度[3,5]。对环境资源利用能力较强的物种,与其他物种往往拥有较高的生态位重叠,但是生态位相似的物种间,资源丰富时可能只是占据相近的生态空间,在生态系统学中的功能和特性不一定相同,对同一空间资源的需求类别存在一定差异,不一定会导致种间竞争[2, 7- 8]。本研究发现,在物种水平上,生态位宽度大的物种间的生态位重叠均较大,而生态位宽度较小的物种间的生态位重叠不一定小,如I类型群落的柽柳和骆驼刺,生态位重叠值为1,这在Sandeep等[39]的研究中也有相同的结果,可能存在着两种原因,一方面物种对生境条件和资源类型的重叠部分较大,在对资源共享的同时也存在竞争关系;另一方面物种在生态系统中的地位需要彼此共同促进而决定其生态学作用和意义。而艾比湖流域属温带荒漠干旱区,资源相对贫缺,具有优势的物种并不会主动割舍资源生态空间,这样在长期适应干旱盐渍化生境过程中,大部分物种对资源匮乏的需求趋于特化而发生趋同适应,生态位重叠普通较高,导致物种间资源竞争激烈,限制了物种共存并驱使群落演替,而群落结构和功能的自身稳定性,反过来又作用于各物种,使其各自扮演着群落中不可或缺的生物生态角色[5- 6]。另外,综合生态位宽度和生态位重叠值的分析可见,群落水平上种群的生态位宽度与生态位重叠具有一定的相关关系,群落类型I和III物种的生态位宽度较大,生态位重叠较高,说明在这2种土壤水盐的环境下,种群间存在较强的竞争关系。群落类型II中物种的数量最多,但是个别物种出现的频率较低,整体上种群的生态位宽度较小,生态位重叠普通较低,物种间的竞争关系相对较弱。这与物种水平上的结果不太一致,可能是因为在资源匮乏的干旱区出现了生境相似的特化种或者伴生种,这些物种生态位宽度值较低,但由于生长或生存的资源纬度相似,物种间生态位重叠值较大,说明生态位宽度和生态位重叠值由物种向群落尺度转换时,两者之间的关系存在冗余,尤其是在生态位重叠值较低时,生态位宽度的冗余效应较大。
3.3 群落生态位对水盐的响应特征
群落生态位宽度可以表示植物群落整体水平对环境资源利用状况尺度的大小,能够反映群落中物种的分布状况,以及间接地衡量群落结构的稳定性,在一定程度上,数值越高表示群落中各物种分布越均匀,该群落的结构越稳定[6]。不同的生境可能会引起群落生态位的变化[7],以往研究表明,自然环境压力会逐渐淘汰掉一些物种,群落的物种丰富度会降低,并且泛化种的数量也会因为生境压力逐渐减少,导致群落的结构组成单一化,那么群落整体的资源利用效率和环境适应能力就会减弱,从而限制群落的生态位幅[6-8]。与之相反,也有学者认为,自然环境压力虽然会对植物进行优劣筛选,但是植物普通的会采用本身的生态学特性、共生或特化等策略保持个体的数量并均匀地分布,提高自身在群落中的生态空间地位[12],群落整体的资源利用能力和环境适应能力会增强,促进群落生态位宽度的增幅[40]。但在该过程中,另有观点认为,在干旱区荒漠基质植物群落中,资源匮乏导致物种间的竞争关系加强,整体群落的生态幅并不会有太大的改变,极端情况下可能会发生抑制作用[4,41]。本研究发现,高水高盐的土壤水盐分配格局对群落的生态位宽度有一定的促进效应,而低水低盐的土壤水盐分配模式对群落的生态位宽度具有一定的限制作用,土壤的水盐环境梯度与群落生态幅呈现一种非线性相关特征,这一结论与上述观点相悖。这可能是因为,近河岸荒漠森林生态系统的资源充足,植物分布均匀,并且能够有效地提高群落中各物种的数量,加强群落的稳定性,群落对资源的利用能力和效率也会有效增加,从而群落生态幅增大[24,40,42],相反,远离河岸的低水低盐土壤环境导致植物分布不均匀,并且群落中各物种的个体数量无法持续维持,群落结构组成单一,稳定性较差,对资源利用的能力和效率整体降低,从而群落生态幅减小[8,35]。另一方面,研究区盐渍化以及干旱缺水的天然缺陷,使水分补给量成为植物生长和群落演替的关键因素,而水分的补给来源和效率在维持植物数量和群落稳定方面可能起着重要作用,近河岸土壤具有高水高盐的分配特征,拥有长期持续性的河水的补给占比较大,容易被植物群落吸收利用,同时水分补给效率也较高,而远离河岸低水低盐的土壤环境,深层地下水的补给方式增加了植物地下的根投入比,其水分补给效率较低[35,39-41],生境间水源可获取的多样性,影响着植物地下和地上的形态结构,进一步改变了群落的生态结构,这个可能是决定研究区植物生态位宽度的重要因素,将成为今后研究的重点。
