柴达木盆地东部荒漠植物生态位特征
2019-05-31牛慧慧张斯琦张博雄
牛慧慧,陈 辉,付 阳,杨 祎,张斯琦,张博雄
河北师范大学资源与环境科学学院,河北省环境演变与生态建设重点实验室, 石家庄 050024
植物生态位研究是近年生态学和地理学研究领域的热点之一[1-5],生态位(ecological niche)不仅可以展现群落种间关系的现实结果,而且是群落特性的发生与发展、种间竞争和进化的动力和原因[6],生态位测度包括生态位宽度和生态位重叠,它们是基于种群在一系列资源状态下的分布数据[7]。生态位宽度是物种所能利用的各种资源的一个综合指标[8]。生态位重叠反映两个物种对同一资源的利用效率及共享程度[9],其重叠值的大小与资源的竞争强弱呈正比。进行生态位研究,定量化描述环境因子是首要一步,现阶段常用的方法是划分资源环境梯度[10],赋以等级值,由此入手计算并分析植物种群生态位,该方法便于探讨不同植物对同种资源环境的利用效率、同一植物对不同资源的利用程度以及物种间的共存关系。目前资源环境因子的选择主要有土壤因子[11- 17],地形因子[2,18- 20],气候因子[21,22]等。生态位对于研究生物群落的种间关系具有普适性,但在现阶段的研究中,研究角度多从不同群落类型或群落演替的不同阶段展开[23- 27],对干旱半干旱区研究主要集中在高寒草地、绿洲过渡带等[28- 31],其中,对荒漠植物的研究主要集中在新疆[14,16,30,32-33]、甘肃等[31,34-35]地的草地或河岸绿洲等地,对青藏高原东北部的柴达木盆地研究较少。
柴达木盆地气候寒冷干燥,人烟稀少,盆地内植被覆盖率小于5%[36],灌木是其主要的优势植物,其群落的稳定性有益于荒漠生态系统的恢复和重建[37],因此以生态位入手准确地评估该地区植物群落的内部关系具有重要生态意义。现阶段针对柴达木盆地这一独特生态地理单元的生态位研究较少,以紫花针茅为代表的天然草地退化方面的生态位研究,天然草地的退化严重影响生态位宽度和生态位重叠的指数[38],对柴达木盆地南缘巴隆地区11种典型植物的生态位研究的结果表明,钝叶猪毛菜是此地区的优势种[39]。柴达木盆地内缺乏植物生态位与生境关系的研究。分析盆地东部典型荒漠植被与土壤环境因子的生态位关系,有助于筛选影响盆地东部植被生长分布的关键土壤环境因子,有助于确定自然条件下本地区的优势植物,使荒漠化防治工作做到有的放矢。故本文拟对柴达木盆地东部13种优势植物在6个土壤因子上的生态位特征进行分析,以期为柴达木盆地的植被恢复和生态保护提供理论参考。
1 研究区概况
研究区位于柴达木盆地东部(图1),95°02′—99°00′E,36°24′—37°37.80′N范围内,海拔2694—4357 m,属于典型的高原大陆性气候,寒冷干旱,降水稀少且多集中在夏季[40],年均降水量从盆地西北部(<15 mm)—盆地中心—盆地东南部(150—200 mm)呈逐渐递增状态,东部湿润程度大于西部[41],盆地内年总辐射量160×4.186—175×4.186kJ/cm2,年均温约在-1.4—5.1℃之间,年蒸发量2000—3000 mm[42]。整个区域风蚀迹地、沙丘、沙漠、盐湖和盐土平原交替出现[43]。盆地东部水平地带上荒漠灌木、小灌木半灌木为主要植被类型,植被相对较为密集的地区主要分布在由高山冰雪融水形成的内流河附近及其派生的湖泊和冲洪击扇等地[44]。在盆地东部边缘有从上部的高寒草甸(小嵩草)过渡为荒漠-草原过渡带(芨芨草)到下部典型荒漠类型的垂直分布[45]。研究区优势植物类型包括:白刺(Nitrariatangutorum)、麻黄(Ephedrasinica)、沙拐枣(Calligogumkozlovi)、柽柳(Tamarixramosissima)、梭梭(Haloxylonammodendron)、驼绒藜(Ceratoideslatens)、盐爪爪(Kalidiumfoliatum)、琵琶柴(Reaumuriasongonica)、蒿叶猪毛菜(Salsolaabrotanoides)、芦苇(Phragmitescommunis)、芨芨草(Achnatherumsplendens)、小嵩草(Kobresia)等13种。

图1 研究区及典型样地位置Fig.1 Research area and typical sample location
2 材料与方法
2.1 样方设置及调查
植物样品采自柴达木盆地东部27个样地(图1),每个样地为同种植物群落,随机调查5—10个样方,不同生活型的植物设置的样方大小亦不同:高寒草甸采用的是0.5 m×0.5 m,其他草本植物采用的都是2 m×2 m的样方,植株较小的灌木群落采用2 m×2 m的样方,植株较大的灌木采用5 m×5 m或10 m×10 m的样方。群落调查内容包括:群落总盖度、种分盖度,植物种类、株高、盖度、频度等。调查样方的同时用GPS定位。
2.2 土样分析
土壤样品采集方法:每个植物样地随机调查3个土壤剖面,深度为0—30 cm,采用机械法每10 cm采集一个样品,每个样品约500 g。分析值为3个剖面所有土层的平均值。土壤含水量采用烘干法测定,土壤容重采用环刀法测定,土壤pH值采用pH试纸,土壤有机质测定采用重铬酸钾容量法,土壤可溶性全盐采用电导法,土壤全氮采用凯氏定氮法。
2.3 研究方法
2.3.1生态位宽度的测度方法
生态位宽度的测定,采用经Corwdl修正的Levins公式,该公式被广泛应用于荒漠群落物种的生态位研究[13,31],生态学意义明确,能够表达群落中优势种生态位宽度的对比关系[46]。
(1)
式中,Bi表示物种i的生态位宽度,代表第i个物种在第j个资源等级下的重要值占该种在所有资源等级中的重要值总和的比例,r表示资源梯度,式中:Pij=nij/Ni,它代表物种i在第j个资源状态下的重要值占该种所有资源等级中的重要值总和的比例[47]。
2.3.2生态位重叠的测度方法
生态位重叠指数采用Pianka(1973)生态位重叠公式计算:
(2)
式中,Oik为物种i和物种k的生态位重叠指数,Pij和Pkj分别为物种i和k在第j个资源等级下的重要值占该物种在所有资源等级上的重要值总和的比例。生态位重叠指数的取值范围为[0,1],当Oik=1时,表示物种i和k在第j个资源等级下完全重叠,而Oik=0时,则说明两个物种不具有共同利用某一资源的状态[48]。本文中重要值的计算采用物种在样方中出现的体积,体积=平均盖度×平均高度,其中平均高度=平均高度和/物种频度/10[49]。
2.3.3资源梯度划分
分析27个样地的土壤因子,结果表明:土壤含水量的变化范围在0.70%—41.99%之间,均值为12.63%,极差高达41.29%;土壤容重变化范围在0.86—1.76 g/cm3之间,均值为1.42 g/cm3,极差达0.90 g/cm3;土壤有机质的变化范围在0.14%—8.23%之间,均值为1.42%,极差达8.09%;土壤pH的变化范围在7.87—9.40之间,均值为8.56,极差达1.53;土壤可溶性全盐的变化范围在155.06—3992.11 EC25μS/cm之间,均值为886.33 EC25μS/cm,极差高达3837.05 EC25μS/cm;土壤全氮的变化范围在0.01%—0.40%之间,均值为0.11%,极差达0.39%。考虑到样地在资源因子梯度上的间距问题[50],为了更准确地表达不同资源维下物种的生态特征,本文对每个土壤指标进行自然对数转换[47],划分12个等级资源梯度(表1)。
数据处理采用Excel 2010和SPSS 19软件。
3 结果与分析
3.1 植物群落调查结果
野外调查共获得170个灌木样方和60个草本样方,累计出现3次以上的共计36种,分属14科36属。其中,以藜科(Chenopodiaceae)、蓼科(Polygonaceae)、柽柳科(Tamaricaceae)、蒺藜科(Zygophyllaceae)、麻黄科(Ephedraceae)、禾本科(Poaceae)、莎草科(Cyperaceae)植物种类居多,其他科仅一种且大都出现在草本样方中。在这些植物中藜科植物占比最高,为20%,是优势科,其他物种分布稀疏且不均,剔除偶见种,本文共涉及7科13种,分别为驼绒藜、沙拐枣、琵琶柴、蒿叶猪毛菜、白刺、麻黄、柽柳、梭梭、合头草、芦苇、芨芨草、小嵩草。
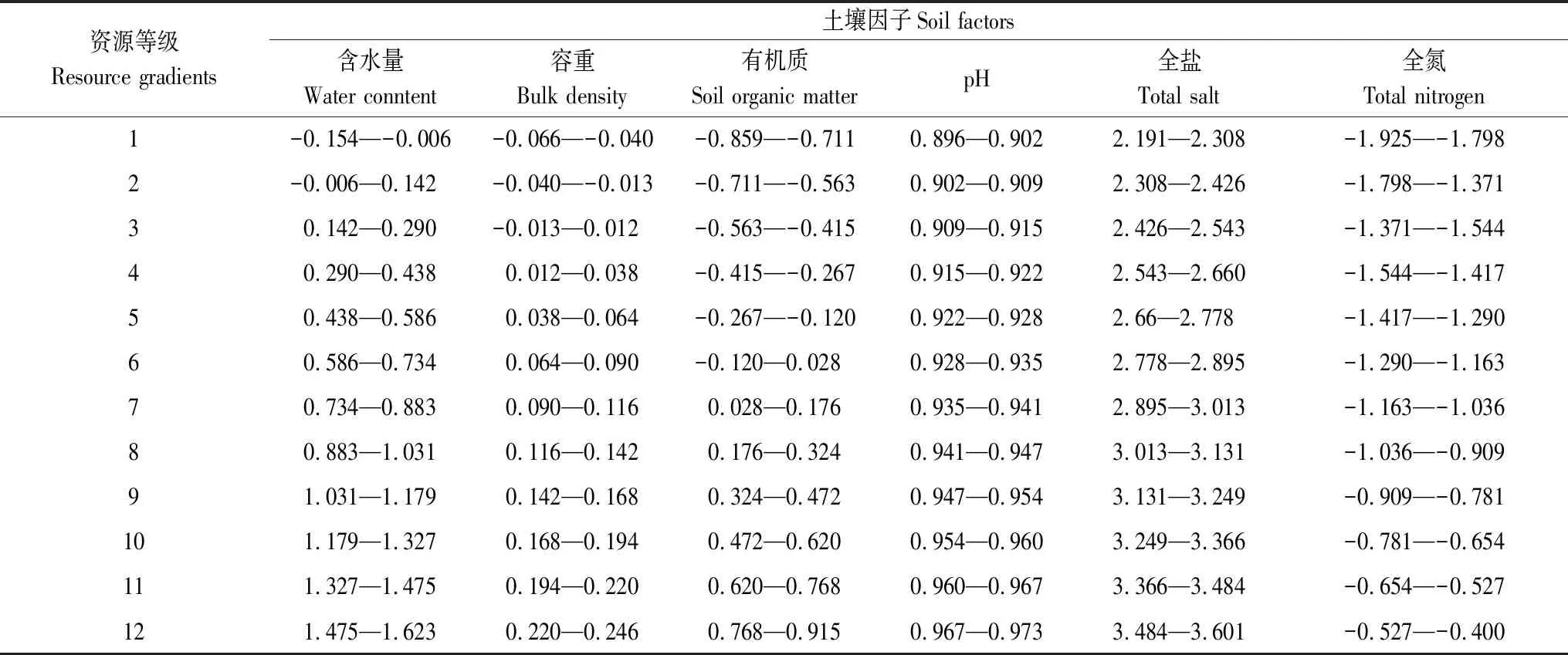
表1 土壤因子资源等级划分
3.2 13种优势植物在6个土壤因子上的分布特征
柴达木盆地生境恶劣,植被组成简单、类型单调、分布稀疏,选取13种出现频率高的植物,分析其在土壤含水量、容重、有机质、pH、可溶性全盐和全氮因子上的生境范围(图2)。在土壤含水量、有机质和全氮因子上,小嵩草皆占据最高值,最低值均出现在沙拐枣物种上,其中,小嵩草在含水量和有机质因子上生境范围最广,芦苇是全氮因子上生存范围最大的物种。与之相反,小嵩草在土壤容重和全盐因子上,均出现在最小值处,其次,在容重因子上占据最大值的是麻黄和沙拐枣,而白刺为容重因子上生存范围最广的物种。土壤含水量与容重呈显著负相关,相关系数r高达-0.985。由此可见,草本植物的土壤生境(较高含水量,较高有机质,较高全氮值)显著优于小灌木的土壤生境(低含水量,低有机质,低全氮)。
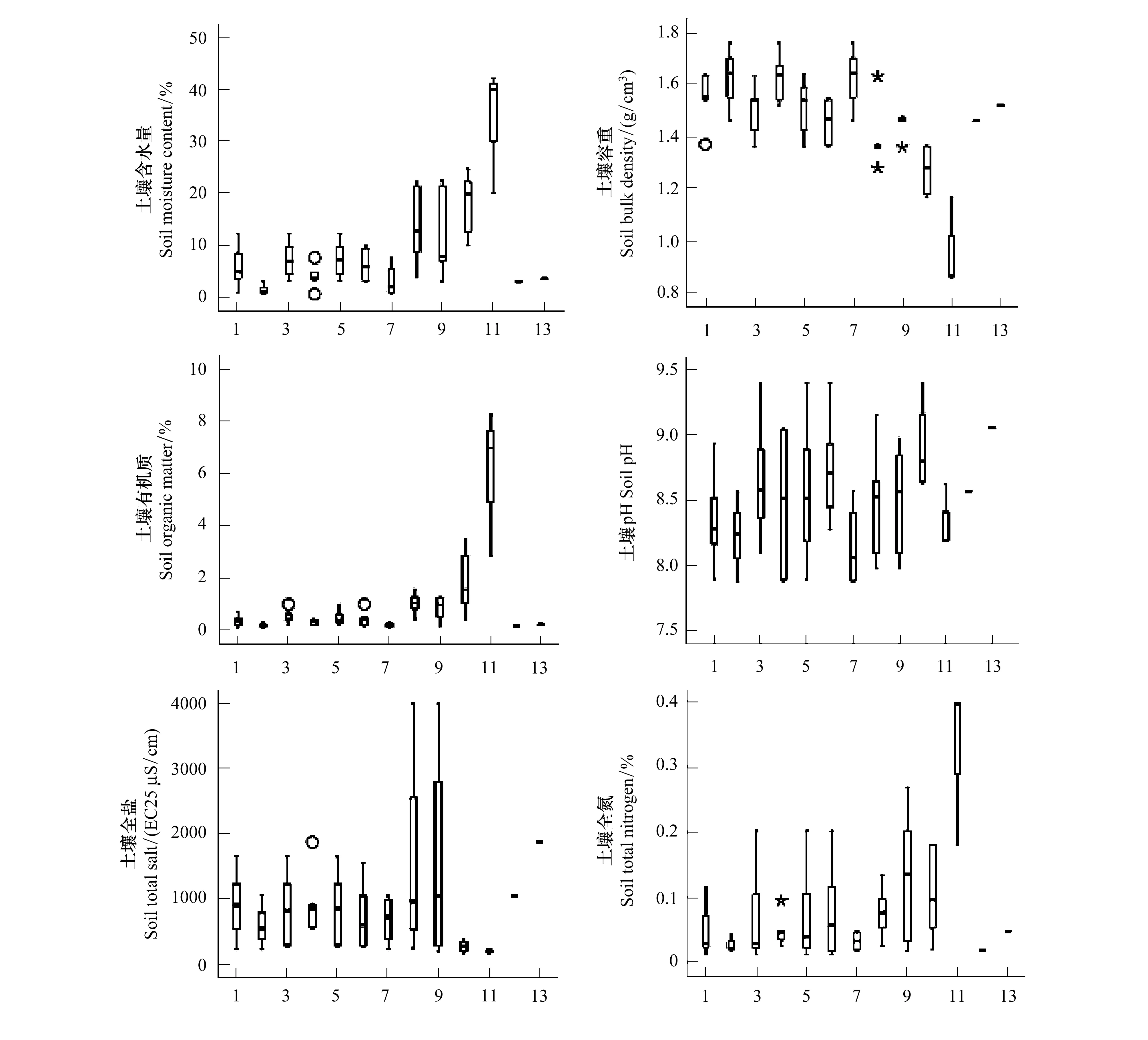
图2 13种植物在土壤因子上的分布特征Fig.2 Distribution characteristics of 13 plants on soil factorsa.土壤含水量,soil moisture content;b.土壤容重,bulk density;c.土壤有机质,organic matter;d.土壤pH, soil pH;e.土壤全盐,total salt;f.土壤全氮,total nitrogen; 横坐标X-axis: 1.驼绒藜,Ceratoides latens;2.沙拐枣,Calligogum kozlovi;3.琵琶柴,Reaumuria songonica;4.合头草,Sympegma regelii;5.蒿叶猪毛菜,Salsola abrotanoides; 6.盐爪爪,Kalidium foliatum; 7.麻黄,Ephedra sinica;8.白刺,Nitraria tangutorum;9.芦苇,Phragmites communis;10.芨芨草,Achnatherumsplendens;11.小嵩草,Kobresia;12.柽柳,Tamarix ramosissima;13.梭梭,Sacsaoul
在全盐因子上,芦苇和白刺明显高于其他物种,盐爪爪、猪毛菜、琵琶柴和沙拐枣也具有较高的盐分适应性,而小嵩草、芨芨草这些草本植物普遍生长在含盐较低的地区,但所有植物生长地土壤盐分含量值均大于150 EC25μS/cm。13种植物在土壤pH因子上的生境相对差异较小,但依然存在高低差值,芨芨草生长在碱性较高(pH值8.6—9.4)的土地上,其他植物的生境地土壤pH也均大于7.9,说明柴达木盆地东部高含盐量土壤和偏碱性土壤广泛分布。
3.3 13种优势植物在6维土壤因子上的生态位宽度
优势物种在6个土壤因子梯度上的生态位宽度计算结果(表2)表明,在土壤水分维度上,驼绒藜的生态位宽度值最大,为0.344,其他生态位宽度值相对较高的物种有琵琶柴、芨芨草、芦苇和合头草,说明驼绒藜、琵琶柴等6种植物对水分的利用能力强。在土壤容重维上,驼绒藜(0.246)和芨芨草(0.222)的生态位宽度值显著大于其他物种,物种盐爪爪在该维上的生态位宽度值是6维中最小的,说明盐爪爪对土壤容重的要求较高。
驼绒藜、芨芨草和琵琶柴在土壤有机质维上具有较高的生态位宽度值,这与其在土壤水分维度上占据较高的生态位宽度值相似。其次,白刺在土壤有机质维上的生态位宽度值明显高于其他5个土壤维度。琵琶柴、驼绒藜、蒿叶猪毛菜和合头草皆具有较高的pH维生态位宽度值。小嵩草在土壤全盐维上生态位宽度值为0.165,但在其他5维上的生态位宽度值均小于0.1,表明小嵩草在盐分维度上与其他12种植物具有相似性。在土壤全氮维度上,琵琶柴的生态位宽度值最高,为0.328,显著高于群落中综合生态位宽度值最大的驼绒藜(0.276),说明琵琶柴对土壤养分含量的要求比驼绒藜低。总而言之,在不同土壤维度下,各物种的生态位宽度差异较大,如芦苇在土壤含水量维上值为0.249,而在有机质维上则较低0.161等。梭梭和柽柳的生态位宽度值比较单一,是因为样方出现次数相对少从而影响生态位宽度计算结果。
此外,计算13种植物在6个土壤因子梯度上的生态位宽度均值,得出物种生态位宽度值大小依次为:驼绒藜、琵琶柴、芨芨草、芦苇、合头草、蒿叶猪毛菜、白刺、盐爪爪、小嵩草、麻黄、沙拐枣、柽柳和梭梭。此结果与这13种植物出现在样方中的频率(图3)具有显著的相关性,相关系数r为0.742。
3.4 13种优势植物在6维土壤因子上的生态位重叠
在土壤水分维度上,柽柳与梭梭的生态位重叠值为1,表明这两个物种在土壤水分生境上完全重合,存在强烈的竞争关系,也有可能是样方截取的原因导致这两种植物完全重叠,暂不确定。其次,柽柳与梭梭除了与合头草(0.797)、驼绒藜(0.600)具有生态位重叠关系外,与其他物种则表现为重叠值极小或完全不重叠的关系。此外,蒿叶猪毛菜与白刺、蒿叶猪毛菜与盐爪爪、麻黄与白刺的生态位重叠值明显高于其他种对,分别为0.937、0.885和0.882。沙拐枣与麻黄的重叠值为0.166,与其他优势种之间的生态位重叠值则较低均小于0.1。在土壤容重维度上,蒿叶猪毛菜与盐爪爪重叠值最大,高达0.994,驼绒藜与琵琶柴、蒿叶猪毛菜、盐爪爪次之,重叠指数分别为0.861、0.741、0.772,这与驼绒藜在土壤水分和容重维上的生态位宽度值最大的结果相契合(表3)。

表2 13种优势植物的生态位宽度
物种编号:1.驼绒藜,Ceratoideslatens;2.沙拐枣,Calligogumkozlovi;3.琵琶柴,Reaumuriasongonica;4.合头草,Sympegmaregelii;5.蒿叶猪毛菜,Salsolaabrotanoides; 6.盐爪爪,Kalidiumfoliatum; 7.麻黄,Ephedrasinica;8.白刺,Nitrariatangutorum;9.芦苇,Phragmitescommunis;10.芨芨草,Achnatherumsplendens;11.小嵩草,Kobresia;12.柽柳,Tamarixramosissima;13.梭梭,Sacsaoul
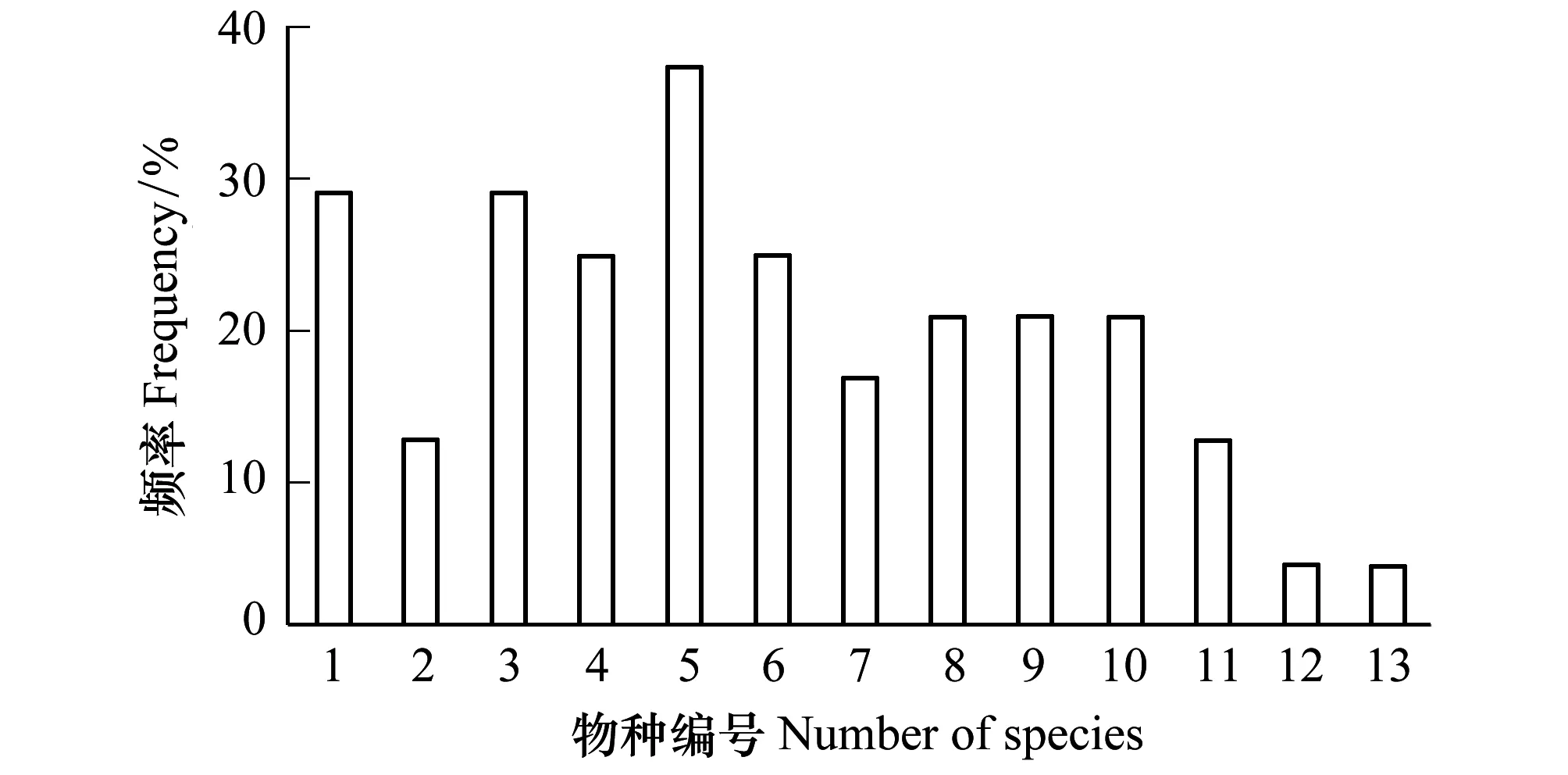
图3 13种优势植物在样方中出现的频率Fig.3 The frequency of 13 dominant plants in the sample横坐标编号含义见图2
在土壤有机质维度上(表4),沙拐枣和柽柳的生态位重叠值高达0.989占据第一位,驼绒藜、琵琶柴与其他物种的生态重叠值相对较高,而麻黄、芦苇与其他物种之间的生态位重叠值较低。合头草与梭梭、白刺与柽柳、琵琶柴与盐爪爪、驼绒藜与琵琶柴在土壤pH维度上的生态位重叠值均较高,分别为0.808、0.947、0.769、0.742。在土壤可溶性全盐维度上重叠值较大的种对有(表5):合头草与麻黄(0.955)、驼绒藜与沙拐枣(0.843)、沙拐枣与芨芨草(0.792)、琵琶柴与盐爪爪(0.779)。在土壤全氮维度上,驼绒藜、琵琶柴、合头草与其他优势种之间的生态位重叠值较高,其中合头草与芦苇之间的重叠值达到0.861。
总的来说,物种生态位宽度越大与其他优势种之间具有重叠关系的可能性越高。纵观物种在6个土壤因子上的生态位重叠值,生态位重叠值<0.1、<0.3和>0.5的物种占总对数的比例,分别表现为含水量维上58%、69%和23%,容重维上55%、73%和21%,有机质维上56%、69%和27%,pH维上68%、86%和10%,全盐维上67%、81%和15%,全氮维上58%、79%和13%,说明物种整体生态位重叠度小,生态位分化显著,尤其是在土壤pH、土壤全盐和土壤全氮上。

表3 13种优势植物在土壤含水量和土壤容重维度上的重叠矩阵
左下角为13种优势植物在土壤水分维度上的生态位重叠矩阵,右上角为13种优势植物在土壤容重维度上的生态位重叠矩阵;物种编号含义见表2。表中“0”为待比较的两种植物其中一种未出现;“0.000”为计算结果不足表中精度
左下角为13种优势植物在土壤有机质维度上的生态位重叠矩阵,右上角为13种优势植物在土壤pH维度上的生态位重叠矩阵;物种编号含义见表2。表中“0”为待比较的两种植物其中一种未出现;“0.000”为计算结果不足表中精度
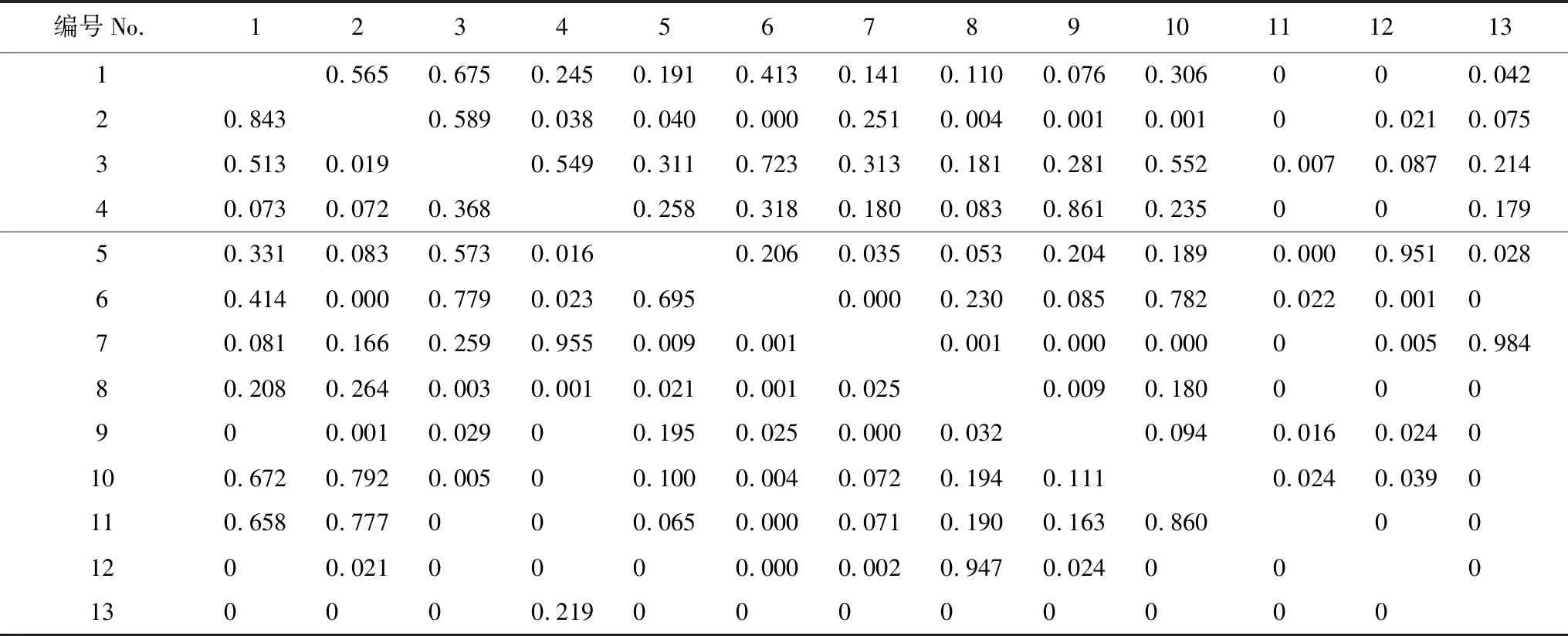
表5 13种优势植物在土壤全盐和土壤全氮维度上的重叠矩阵
左下角为物种在土壤可溶性全盐维度上的生态位重叠矩阵,右上角为物种在土壤全氮维度上的生态位重叠矩阵。物种编号含义见表2;表中“0”为待比较的两种植物其中一种未出现;“0.000”为计算结果不足表中精度
4 讨论和结论
4.1 讨论
生态位宽度是物种所能利用的各种资源的一个综合指标,生态位宽度大,意味着物种在资源上利用不挑剔或者对某一资源有偏好,且在数量上要求不高;生态位宽度小,意味着该种对资源的质和量要求严格或狭窄[8]。本文以修正的Levins生态位宽度公式计算柴达木盆地东部13种优势植物得出,在6种土壤资源因子上,驼绒藜和琵琶柴均占据较高的生态位宽度,说明这两种植物对土壤因子的质和量要求均比其他植物低,对研究区的环境适应性好,也反映该两种植物在一定程度上地理分布比较均匀[51],倾向于发展为泛化物种。而柽柳、梭梭、麻黄和小嵩草在6个土壤资源上的生态位宽度都比较小,说明这些物种对资源的利用具有局限性,仅能适应独特的生境,在研究区内分布稀疏,以上结论与实际群落样方调查结果吻合度较高[49]。
生态位重叠反映两个物种对同一资源的利用效率及共享程度[9],其重叠值的大小与资源的竞争强弱呈正比。众多研究结果表明,生态位宽度与生态位重叠具有正相关关系[18,29]。本文利用Pianka生态位重叠公式计测结果显示,生态位宽度较大的物种与其他优势种之间的生态位重叠普遍较高,但其在每个土壤因子上并未占据最高值,而是介于中游位置,最高生态位重叠均出现在生态位宽度较小的物种上,如盐爪爪和芦苇在土壤有机质因子上重叠值高达0.957,白刺和柽柳在土壤pH上重叠值达0.947,合头草和麻黄在土壤可溶性全盐因子上重叠值为0.955,麻黄和柽柳在土壤全氮维上重叠值高达0.984。这一现象与张伟等[32]对新疆伊犁荒漠草原生态位的研究,张继义等[26]对科尔沁沙地的生态位重叠研究结果相似。究其原因,一方面是这些物种对资源汲取相似,对土壤生境要求接近。另一方面可能是研究区植物分布稀疏不均,小范围采样中优势种空间分布相近,使得结果出现某一资源维上高度重叠,该误差是否会产生较大的影响还需进一步研究。从竞争的角度看,有的学者认为重叠度高意味着竞争激烈,也有研究表明,高重叠度并不一定会导致竞争。本文所有种对在每一个土壤因子上的生态位重叠值中<0.3的对数占总对数的比例高达76%,说明物种之间生态位重叠总体较低,对资源利用的相似性较小,这表明柴达木盆地东部植物生态位分化显著,整体竞争较小,处于一个相对稳定的状态[49]。
考虑到柴达木盆地地理位置的特殊性,对于植被的恢复和重建应以自然恢复为主,人工辅导为辅的指导理念。盆地东部植被恢复中小灌木可选择驼绒藜和琵琶柴,这两种植物在研究区内环境适应力强,在土壤含盐量高的区域可选耐盐性高的白刺和芦苇,选择适宜生境的植物种类,可能省略不必要的人工的维护,即可融入自然,而后繁衍形成群落。
4.2 结论
本文以柴达木盆地东部27个典型样地所采集的植物和土壤为研究对象,利用Levins生态位宽度公式和Pianka生态位重叠公式计算了13种优势植物在土壤含水量、容重、有机质、pH、全盐以及全氮维上的生态位特征,结果表明:
(1)13种植物在6个土壤因子上平均生态位宽度从大到小依次是:驼绒藜、琵琶柴、芨芨草、芦苇、合头草、蒿叶猪毛菜、白刺、盐爪爪、小嵩草,其中,驼绒藜在土壤含水量、容重和有机质上占据最高值,琵琶柴在土壤pH、全盐、全氮维上占据最高值。
(2)生态位宽度较大的物种在生态位重叠上并不占据最高重叠值,而是处于一个中游的位置。驼绒藜和琵琶柴在生态位宽度和生态位重叠上均占相对高的位置,对群落的建群起重要作用。
(3)78个种对在每一个土壤因子上的生态位重叠值<0.3的占总对数的比例为76%,柴达木盆地东部植物总体生态位重叠偏低,竞争较小,群落处于一个相对稳定的状态。
本研究从6个土壤因子的角度分析了柴达木盆地东部植物的生态特征,未考虑气候因子以及小生境范围内地形等因素对植物生态分布的影响,同时荒漠区植物自身的生活型也未作深度分析,下一步计划加入上述因子,进一步深化柴达木盆地荒漠植物的研究。
