放牧干扰下短花针茅植丛对表层土壤性质的影响
2019-05-22杨新国古君龙杨东东
苗 翻, 杨新国, 古君龙, 杨东东
(1.宁夏大学 西北土地退化与生态恢复国家重点实验室培育基地, 银川 750021;2.宁夏大学 退化生态系统恢复与重建教育部重点实验室, 银川 750021;3.宁夏大学 西部生态与生物资源开发联合研究中心, 银川 750021)
短花针茅(Stipabreviflora)草地是宁夏盐池县荒漠草原地区一种重要的放牧草场类型,在放牧干扰情况下,往往面临着比较严重的退化压力,包括植丛小型化,覆盖度和连续度的下降[1],土壤风蚀退化过程加强等[2]。表层土壤轻度沙化、短花针茅植丛空间分布破碎化是研究区短花针茅草地的一种典型的早期退化现象。在发生不可逆的严重沙化之前,破碎化点状分布的短花针茅植丛与轻度沙化的土壤空间格局的相互作用,可能是影响草地在放牧干扰下系统稳定性自我维持的一种关键生态学过程[3-4]。目前,我们对这种小尺度上植被与土壤相互作用的认识还不够深入。
放牧干扰通常会加速生境破碎化,增大土壤异质性;围封则会促进草地植被恢复,降低土壤异质性[5]。在干旱半干旱区,土壤养分与细颗粒物质往往在灌丛下富集,引起明显的“肥岛”现象,尤其表现在土壤表层[6-7]。相关研究显示:植物的演替与土壤性状的变化存在互馈效应,进而改变土壤颗粒组分与养分物质的空间分布关系[8],植物根系的分布状况也会显著影响土壤资源的分布[9],有根系的地方明显可以产生优势流,有利于土壤水分和养分物质聚集迁移[6],根区优势流往往导致土壤颗粒与养分物质的再分配。韩凤鹏等通过比较有根系和无根系的长芒草植被邻近坡面发现,植丛根系可以提高土壤养分的运移,对土壤物理化学性质影响非常明显,特别是对0—20 cm土层[9]。因此,小尺度上土壤和植被的相互作用,特别是由于植丛本身及其根系分布对土壤物质再分布的影响,可能是放牧干扰下草地稳定性维持的一个重要途径,但是由于植物类型、退化程度、生境条件的不同,这种作用的生态后果也不尽相同。本研究以放牧干扰下退化短花针茅草地为研究对象,以临近的围封恢复草地为对照(CK),对比研究放牧干扰下短花针茅植丛边缘以及丛外裸斑表层土壤颗粒组分和养分特征,分析其小尺度空间分布规律,及其相关关系在空间上的变化。旨在阐明:(1) 短花针茅植丛是否会起到局部抑制土壤退化的作用?(2) 在放牧干扰对上述生态过程产生何种影响?进而深化对小尺度植被与土壤相互作用格局与机制的认识,为荒漠草原早期退化短花针茅草地的优化管理提供理论依据。
1 研究区域与研究方法
1.1 研究区概况
研究区位于宁夏回族自治区盐池荒漠草原,北与毛乌素沙漠相连,南靠黄土高原,属于典型的草原向荒漠草原的过渡地带。受地理位置和气候条件影响,该区域植被多样性低,土壤严重贫瘠,生态系统脆弱较不稳定。本区气候属于典型的中温带大陆性气候,年平均气温-8℃左右,最热和最冷月是7月(平均气温22℃左右)和1月(平均气温于8℃左右),年日照时数2 860 h,年均无霜期165 d,年均降水量280 mm,降雨量年际变率大,60%左右的降雨集中在每年的7—9月,年潜在蒸发量处于2 700 mm左右。地带性土壤主要以灰钙土(淡灰钙土)为主,非地带性土壤主要有风沙土和草甸土等。该区植被类型以旱生和中旱生类型的草本和沙地小半灌木植被为主,主要优势植物种有苦豆子(Sophoraalopecuroides)、中亚白草(Penni-setumcentrasiaticum)、猪毛蒿(Artemisiascoparia)、牛枝子(Lespedezapotaninii)、草木樨状黄芪(Astragalusmelilotoides)和阿尔泰狗娃花(Heteropappusaltaicus)等。
1.2 选择样地与样品采集
1.2.1 样地选择 试验样地位于盐池县杨寨子村(37°49′N,107°27′E),选取以短花针茅为优势种的自由放牧草地为研究对象,以围栏封育为对照,构成本研究的对比样地体系。围栏封育前,两类试验样地都处于自由放牧状态(1.5只羊/hm2),放牧强度相对均一,在当地属于重度放牧水平。围栏封育始于2003年,之后没有任何的放牧干扰,植被和土壤得以持续恢复。围栏外则一直维持放牧利用,表层土壤沙化明显,植被低矮稀疏。两类试验样地内各设置1个50 m×50 m的标准样地作为试验调查区,在标准样地内各随机选取5个1 m×1 m的样方(总共10个样方),调查样地短花针茅群落的基本植被情况(表1)。

表1 围封区和放牧区的短花针茅群落植被情况(均值±标准误)
注:短花针茅是一种密丛型植物。
1.2.2 土壤样品采集 2017年9月在每个随机选取的10个植被调查样方内,随机选定1丛短花针茅为对象,分别在植丛边缘与丛外裸斑对表层0—14 cm深度范围的土壤进行分层采样,分层深度为0—2 cm,2—4 cm,4—6 cm,6—8 cm,8—10 cm,10—12 cm和12—14 cm共7层。土壤分层采样使用特制的一体化分层土壤采集器完成(图1)。该设备能够一次完成所有分层土壤的采样,提高了野外采样的效率;同时避免逐次分层采样时手工挖取土壤造成的误差,保证了精细分层采样时不同样点之间深度的一致性。具体的采集步骤,首先沿植丛南北轴线和10 cm外丛外裸斑处挖两个深20 cm,宽20 cm,长50 cm的作业槽,修整作业槽侧壁的土壤断面,分别将土壤采集器打入作业槽侧壁,在作业槽南北两侧端面各取一次样,并分层混合两侧土样装袋,分别为植丛边缘和丛外裸斑每2 cm一层的土样。总共采集到140个土样。
1.2.3 土壤样品分析以及数据测定方法 采集的140个土壤样本带回实验室阴干剔除可见的动、植物残体和石块后,分别进行土壤颗粒组分和土壤养分(有机碳和全氮)的测定。土壤颗粒组分(PSD)使用英国马尔文公司的Mastersizer 3000激光衍射粒度分析仪进行测定,该型号粒度仪可自动测定土壤粒径的体积百分比,粒度分析的粒径范围0.01~3 500 μm,重复性误差±0.5%,准确性误差≤±1%;测定前土样过2 mm的筛子,不需要进行预处理,直接上机测定;土壤颗粒组分分级采用美国制土壤颗粒组分分级标准,分为黏粒(<2 um)、粉粒(2~50 um)、砂粒(50~2 000 μm)[10-11]。土壤有机碳(SOC)使用日本岛津TOC-L有机碳分析仪ssm-5000年固体模块测定,该仪器采用680℃燃烧催化氧化法,测量范围4 μg/L~30 000 mg/L;土壤全氮(TN)使用德国艾力蒙塔(vario MACRO cube )元素分析仪进行测定。土壤养分测定前运用球磨机研磨使其全部可以通过150目的筛子,土壤有机碳(SOC)称量30 mg左右,土壤全氮(TN)称量150 mg左右制样,然后上机自动完成测定。
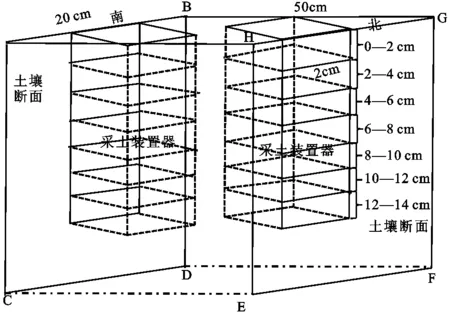
注:带虚线面为封闭面,面ABCD和面EFGH为土壤断面刀口面。
图1 一体化分层土壤采集器示意图
1.3 数据处理分析
所得试验数据统一用Office Excel 2010进行数据录制整理及图表制作,采用SPSS 22.00统计软件中的One-Way ANOVA和Peareson 相关分析方法分别进行方差分析和相关性分析。
植物群落计算其多样性指数:计算物种重要值,Shannon-Wiener多样性指数、Pielou均匀度指数。各指数计算公式[12]如下:
物种的重要值(Ⅳ)=(相对盖度+相对密度+相对频度)/3
Shannon-Wiener多样性指数(H1):
H1=-∑(pi)lnpi
Pielou均匀度指数(E1):
E1=H1/lnS
2 结果与分析
2.1 整体土层土壤颗粒组分、有机碳(SOC)和全氮(TN)的分异规律
分层土样全部混合所得土样(0-14 cm)的测定结果反映了整体土层的变化规律。如图2所示,因生境不同土壤颗粒组分围封区和放牧区存在显著差异(p<0.05),无论植丛边缘还是丛外裸斑,围封区的土壤黏粒含量(45.95%)远高于放牧区(29.06%),相应地砂粒含量则是围封区(29.07%)低于放牧区(42.46%);围封使植丛边缘土壤黏粒含量显著增高了16.69%,粉粒和砂粒显著降低了3.50%,13.40%(p<0.05)。植丛边缘与丛外裸斑无显著差异(p>0.05)。
土壤SOC与土壤黏粒含量表现出相同的变化规律,围封区和放牧区植丛边缘的土壤SOC平均含量分别为0.72%和0.57%,围封显著提高了表层土壤SOC含量(p<0.05),植丛边缘与丛外裸斑无显著差异(p>0.05)。土壤TN在各种生境之间都无显著差异(p>0.05),具有全局的均一性和稳定性。
2.2 精细分层下土壤颗粒组分、有机碳(SOC)、全氮(TN)的分异规律
精细分层土样的测定结果不但反映了不同生境之间土壤的分异规律,还进一步反映了其影响因素的作用深度。如图3所示,无论植丛边缘与丛外裸斑,围封区上部0—12 cm土层黏粒含量显著高于放牧区,砂粒含量显著低于放牧区(p<0.05);黏粒和砂粒仅在12—14 cm最下土层无显著差异(p>0.05);而粉粒在0—8 cm土层受围封影响而显著提高(p<0.05),8—14 cm土层无显著差异(p>0.05)。在放牧区和围封区,植丛边缘与丛外裸斑之间均无显著差异(p>0.05)。围封区植丛边缘处0—2 cm,2—4 cm和12—14 cm土层的SOC显著高于丛外裸斑(p<0.05),但是放牧区植丛边缘与丛外裸斑依然无显著差异(p>0.05)。
同一生境下不同深度土样之间的比较反映了土壤颗粒组分的垂直分异规律。如图3所示,就围封区而言,植丛边缘与丛外裸斑的垂直分异规律基本一致,没有明显的突变层。黏粒含量表现出随着土层加深而增大的趋势,变化范围为39.98%~50.60%,但在相邻土层之间无显著差异(p>0.05);对放牧区而言,黏粒随土层深度的变化略有不同,表现出一种突变模式:在0—10 cm上部土层基本保持在26.23%~26.83%,随后陡然增高,在10—12 cm和12—14 cm 显著提高到31.32%和40.2%(p<0.05)。砂粒的变化趋势与黏粒相反,随土层加深而递减。粉粒含量的变化趋势与砂粒相似,随土层加深递减,但变化的深度和范围均小于砂粒。围封区的变化集中在0—6 cm土层,变化范围为28.34%~24.56%;在放牧区呈现出持续降低的趋势,变化范围为32.23%~22.55%。

注:图中有4种生境:围封区植丛边缘、围封区丛外裸斑、放牧区植丛边缘和放牧区丛外裸斑;土壤颗粒组分体积百分含量(大写字母表示不同生境条件下同一级颗粒组分的显著性差异(p<0.05),图中有机碳和全氮(大写字母表示不同生境处理下的含量显著性差异(p<0.05))。
图2 不同级土壤颗粒组分、有机碳和全氮百分含量分布特征
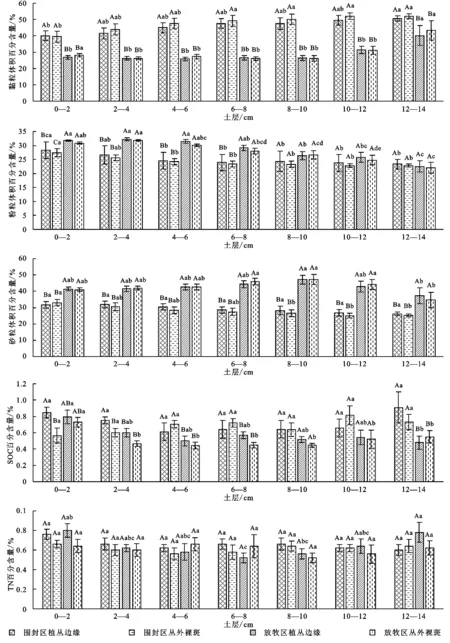
注:图中有4种生境:围封区植丛边缘、围封区丛外裸斑、放牧区植丛边缘和放牧区丛外裸斑;土壤粒径体积百分含量(大写字母表示4种不同生境条件下同一级粒径的显著性差异(p<0.05),图中有机碳和全氮(大写字母表示不同生境处理下的含量显著性差异(p<0.05));小写字母表示同一生境条件下随土层加深含量变化的显著性差异(p<0.05)。
图3 土壤颗粒组分、有机碳(SOC)和全氮(TN)的垂直分布特征
围封区SOC和TN随土层加深的变化都不显著(p>0.05),但放牧区0—2 cm土层SOC表聚性突出,含量为0.79%,显著高于其他土层(P<0.05),TN在植丛边缘呈现“V”形变化,在6—8 cm土层的含量仅为0.05%,显著低于其他土层(p<0.05)。
2.3 土壤颗粒组分、有机碳(SOC)、全氮(TN)之间的相关关系
2.3.1 整体土层 对不同生境下土壤颗粒组分之间,及其与土壤SOC,TN之间进行了相关分析结果详见表2。首先,围封或放牧显著影响着土壤颗粒组分的内部构成关系。围封条件下黏粒、粉粒与砂粒之间普遍存在极显著的负相关关系(p<0.01),但在放牧条件下,粉粒与黏粒、砂粒的相关性丧失,仅黏粒和砂粒仍保持极显著的负相关关系(p<0.01),但是相对有所弱化。无论放牧还是围封条件下,植丛的存在都没有影响上述关系。其次,围封区颗粒组分与SOC,TN的相关性都不显著(p>0.05),放牧区只有植丛边缘的TN与黏粒极显著的正相关关系,丛外裸斑的SOC与砂粒表现出极显著的负相关关系(p<0.05)。

表2 不分层条件下颗粒组分和有机碳、全氮的相关分析
注:*为p<0.05显著水平,**为p<0.01显著水平。
2.3.2 分层土壤剖面 由表3看出,颗粒组分三大分级组分间的相关系数(R)值因生境不同而表现出完全不同的变化趋势。围封区不同级颗粒组分的相关关系随土层深度的变化基本稳定,特别是丛外裸斑处上下通体几乎没有任何变化,只是植丛边缘的粉粒和砂粒的(R)值在0—2 cm和8—10 cm土层出现了两个明显的突变点,显著性丧失。放牧区在12—14 cm土层黏粒和粉粒、粉粒和砂粒发生相关关系的逆转,导致其垂直分布趋势出现了明显的变化;砂粒和黏粒的显著负相关关系分布格局与围封区基本一致,但下土层的变异性明显加大。另外,放牧区黏粒和粉粒的(R)值显著相关集中在2—8 cm土层,粉粒和砂粒集中在2—10 cm,0—2 cm和12—14 cm土层。所以,近地表的外因扰动和下层土壤性质的变化改变了土壤内部不同颗粒组分的构成关系,从而呈现出随土层深度变化不同的垂直变化趋势。
颗粒组分与SOC的相关性一定程度上反映了SOC的形成与保存的土壤环境机制。首先,颗粒组分与SOC的相关性在垂直土壤剖面的分布变化不明显,通体基本稳定,生境因子影响不明显。其次显著相关集中出现在放牧区10 cm以下土层,包括10—12 cm土层黏粒和SOC的正相关,以及10—14 cm土层砂粒和SOC的负相关。结合颗粒组分和SOC绝对含量的变化,整体而言,SOC和黏粒或砂粒在土壤中的形成分布是两个相对独立的过程。
就表(4) 不同级土壤颗粒组成与TN的R值:相对与SOC和颗粒组分的相关关系垂直分布规律,TN表现出了更大的变异性。围封区2—8 cm土层植丛边缘与丛外裸斑的土壤颗粒组分和TN的相关性呈现一种近乎相反的变化趋势,植丛边缘处的负相关和丛外裸斑的正相关,尽管这种相关性并不显著(p>0.05);放牧区植丛边缘与丛外裸斑相关性趋势基本一致,只是在个别土层出现了突变点。显著相关主要集中在放牧区,包括6—14 cm土层砂粒和TN的显著负相关(P<0.05),8—10 cm土层粉粒和TN的及显著正相关(P<0.01),以及10—12 cm土层黏粒和TN的显著正相关(p<0.05)。就与颗粒组分的相关性而言,TN的表现要好于SOC,说明TN的潜在来源与颗粒组成的相关性更高,而且这种关系主要针对的是下层受环境干扰较少的原生土壤。

表4 分层条件下颗粒组分和有机碳、全氮的相关分析
注:*为p<0.05显著水平,**为p<0.01显著水平。
3 讨 论
过度放牧是干旱半干旱草地沙漠化的主要原因之一。放牧不仅影响草地植被的物种组成、群落结构和生产力水平,同时也对草地生态系统的土壤物理结构与化学性状产生影响[13],是荒漠草原土壤颗粒组分和养分分布改变的主要因素之一[14]。国内外学者研究认为,在重度放牧强度下放牧通常会加大家畜对植被的干扰破坏,草地植被随家畜啃食而减少;相比于围栏封育地上植被覆盖度、多度大大降低,地表的裸露程度增大,加速生境破碎化[5]。地表的裸露加大了风沙活动,风蚀作用的加强使得土壤表层细颗粒(黏粒(<0.01 μm))组分降低,促使土壤粗颗粒(砂粒(50<2 000 μm))储存下来,进而改变土壤颗粒组分分布[15-16]。本研究中放牧干扰显著降低了土壤黏粒组分,砂粒显著提高(p<0.05)(图1),与前人研究结论基本一致[15]。土壤养分的变化是一个相对复杂的动态变化过程。国内外学者针对放牧对土壤中养分(SOC和TN)含量的影响研究,由于研究对象、放牧管理方法、时间等方面的差异,所得到的结果不尽一致[17]。相关研究认为放牧对土壤SOC的影响受到植被和土壤的初始状况、环境因素(水分和温度)以及放牧情况(强度、频率、持续时间和动物类型)等综合因素的作用[18]。有些学者认为草原生态系统具有一定的弹性,适度放牧对土壤SOC无显著影响[19],有些则认为放牧显著降低荒漠草原土壤SOC含量[20]。Frank等[21]发现与围封禁牧相比,重度放牧并未使土壤SOC降低,其原因可能是因为食草动物排泄物的归还使土壤表层速效养分增加,土壤的矿化作用加强。本研究中相比于围栏封育,放牧干扰下土壤SOC显著降低,土壤SOC的降低与土壤黏粒组分的降低趋势相同。产生上述现象的原因是因为重度放牧使草地初级生产固定碳素的能力降低,并且家畜的采食凋落物和枯落物减少导致有机碳素由植物凋落物向土壤中输入的减少,从而降低了土壤SOC的来源。家畜的过度践踏破坏了土壤的物理性状,促进草地土壤的呼吸作用,加速碳素由土壤向大气释放[22],减少了土壤中的SOC储量。本研究关于放牧对土壤中TN含量对放牧的响应,发现土壤TN对生境环境的响应并不敏感(图2),具有全局的稳定性和均一性,重牧土壤TN并未降低。上述结果与安慧等[17]在2013年对宁夏盐池县荒漠草原的研究并不一致。原因或许与研究区放牧时间和管理方法的差异有关。一般而言,土壤黏粒分别与土壤SOC和TN呈显著正相关关系,土壤黏粒组分的流失往往导致土壤SOC和TN的下降[16,23-24],这与本研究结果有所不同。这或许表明在本研究(0—14 cm)土层的空间尺度上土壤TN的分布与变化可能是一个相对独立的过程,短花针茅植丛作用下表层土壤质地存在二次过程,受其他环境因子的影响更大,或者说土壤SOC和土壤TN在某种程度上的潜在来源存在明显的区别,所以两者与黏粒的相关关系的垂直变化趋势也不尽相同。
植丛的存在是荒漠草原土壤沙漠化抑制的主要因素之一,显著影响着地表以及地下生物、非生物资源的再分配[25]。在荒漠草原区植丛具有独特斑块状分布特征[26],不同植物土壤养分的空间分布存在明显的异质性,通常会发生明显的“肥岛”现象,随植物中心向外逐渐降低[27]。本研究通过对重度放牧干扰下短花针茅植丛边缘与丛间裸斑的土壤颗粒组分与养分(SOC和TN)分布的比较,发现在此环境下短花针茅植丛边缘与丛外裸斑土壤颗粒组分、养分(SOC和TN)含量无显著差异(表2)(p>0.05),“肥岛”效应反馈并不明显。相对于灌丛的“肥岛”效应,短花针茅植丛“肥岛”效应并不明显,植丛的存在并没有起到局部抑制草地土壤退化的作用,植丛对潜在的风蚀物质的拦截富集作用并不明显[15,29]。与张生楹等对高寒牧区醉马草草丛肥岛效应的研究结果不太一致[28]。主要原因由于植被类型、生境条件以及研究区植丛边缘与丛外裸斑的距离和研究土层深度的不同所导致植物对土壤性质的影响不同。或者说在放牧干扰下短花针茅植丛的“肥岛”效应可能存在更广、更深的空间范围,但具体原因可以考虑加大植丛边缘与丛间裸斑的距离以及土层深度的研究。
荒漠草原由于放牧对植被的直接作用,间接导致土壤理化性质的改变,不同土层土壤颗粒组分和养分(SOC和TN)有所差异。相关研究所示不同土壤利用下土层剖面各土层土壤母质来源和质地有所差异,呈现不同分布特点。在长期的放牧干扰和围栏封育土壤利用下,不仅改变了土壤的自然环境形成条件,同时影响着土壤颗粒组分、土壤SOC的含量及其他理化指标含量分布,各指标性质间的相互关系也随之改变[30]。本研究中放牧干扰下随土层加深植丛的存在仅TN 变化趋势有所差异,其余颗粒组分和SOC都没有显著差异,颗粒组分与养分的显著相关性出现于放牧干扰下。表明在此放牧干扰下土壤颗粒组分与养分的分布关系更为密切,短花针茅植丛的存在对不同土层SOC和TN的影响不同,而TN的分布与变化或许是一个相对独立的过程,影响因素更为复杂。相比于围栏封育,颗粒组分分布的变异从10 cm土层以下开始,SOC(0—2 cm)表聚型聚集,TN则在植丛边缘“V”型(6—8 cm)分布特征,先降低后增高的趋势。这是由于本研究在重度放牧干扰下10 cm土层以下的土壤颗粒组分含量基本不受外在作用力的影响,而是受稳定原生母质来源的影响[30]。本研究于7月份,植物生长最为茂盛,风蚀作用相比于其他月份较小,土壤表层粪便中有机质的转换使得表层0—2 cm土层SOC含量增加。
由此可见,研究区的气候环境条件、植丛种类及其空间分布格局等因素均会导致草地土壤性质间的差异,小尺度植被和土壤间的互馈关系是多种环境因子交互作用的产物,全面揭示其形成机理,以及探讨其对干旱与半干旱生态系统结构、功能与动态的影响,仍需要更系统研究。
4 结 论
(1) 黏粒(<2 μm)与砂粒(50~2 000 μm)的分布呈现相反趋势,有机碳(SOC)与黏粒的分布变化规律一致,呈正相关关系,砂粒则呈负相关关系。放牧干扰显著降低了整体土壤黏粒和SOC含量,对土壤TN无显著影响。相比于围封区,放牧干扰下土壤黏粒和粉粒含量在土层底层会出现突变型增大或者较小;土壤颗粒组分与养分(SOC和TN)的再分布基本是两个相对独立的过程。
(2) 放牧干扰下植丛的存在并没有起到局部抑制土壤退化的作用,土壤SOC反而是围封区植丛边缘处浅层和下层SOC含量相对提高;围栏封育确实推动了土壤黏粒的下行富集。
