粟酒裂殖酵母中ppr基因缺失引起细胞絮凝的机制研究
2019-05-16张娟
张 娟
(南华大学 生药学与生物科学学院,湖南 衡阳 421001)
酵母的无性絮凝是一个可逆的、钙离子依赖性的过程[1]。当絮凝现象发生时,成百上千的细胞会相互吸附到一起,形成细胞团并沉到培养基的底部[2]。引起酵母发生絮凝的内外因素有很多,如营养不足[3-5]或环境中存在有害刺激时[6],单个酵母细胞会聚集到一起形成生物膜用以抵抗外界刺激对整个群体的危害。至于内部因素,有报道指出一旦某些基因的突变、缺失或高表达会诱导细胞出现絮凝现象[7-8]。如酿酒用酵母的絮凝功能对发酵工业就很重要,因为它可以在发酵结束时为生产者提供一个高效、环保、简单、低成本的将酵母细胞和发酵产品分开的方法[9]。已有研究表明,在粟酒裂殖酵母中絮凝可能受到转录因子和编码絮凝蛋白基因及细胞壁重构酶基因的多重调控[10]。粟酒裂殖酵母中至少包含9个絮凝基因(gsf2/pfl1和pfl2~pfl9)。其中,gsf2是编码絮凝蛋白结构域所必需的,pfl2~pfl9则是非必需的[10]。单独高表达pfl基因对引发絮凝非常重要[10]。gsf2高表达会使絮凝程度更严重,但是pfl9高表达不会出现絮凝程度加深的表型[10]。在粟酒裂殖酵母中涉及絮凝的转录因子有Cbf12、Mbx2、Cbf11和Rfl1。Cbf12和Mbx2起转录激活作用,而Rfl1和Cbf11起转录抑制作用[10-13]。与芽殖酵母中絮凝不同的是粟酒裂殖酵母中的絮凝可以被半乳糖抑制,但是不能被甘露糖、葡萄糖和蔗糖所抑制。这个不同反映出两种酵母细胞壁结构存在差异。到目前为止,在粟酒裂殖酵母中的10个PPR蛋白已经全部被鉴定。PPR1~PPR10在线粒体基因表达过程中发挥重要作用[14-15]。Ppr9 (Rpo41) 是线粒体RNA聚合酶;Ppr1的作用主要是特异性稳定cox2和cox3的mRNA;Ppr2 影响线粒体蛋白的翻译;Ppr3在细胞内的主要作用是稳定线粒体中的小rRNA,与芽殖酵母中的Dmr1[16]和人体中的PTCD3[17]具有功能同源性;Ppr4是cox1mRNA特异性的翻译激活因子;Ppr5是线粒体基因的负调控因子,Ppr6与芽殖酵母中的Atp13/Aep2序列同源,特异性影响atp9mRNA的稳定性;Ppr7影响atp6mRNA的稳定性;Ppr8对线粒体基因的翻译有着微弱的影响[18];Ppr10是一个线粒体翻译因子。另外,ppr3、ppr4、ppr6和ppr10的缺失会扰乱细胞内铁平衡,引起细胞调亡[19]。在前期实验中,将野生型菌株和Δppr1、Δppr2、Δppr3、Δppr4、Δppr5、Δppr6、Δppr7、Δppr8和Δppr10置于三角瓶中培养至稳定期,取少量菌液制成装片放置于显微镜下观察,发现Δppr3、Δppr4、Δppr6和Δppr10会引起细胞凝集。因此选择Δppr3、Δppr4、Δppr6以及Δppr10继续进一步的研究。
在芽殖酵母中细胞絮凝的机制研究已经较为成熟,但粟酒裂殖酵母中ppr基因缺失引起细胞絮凝的机制还不清楚。因此,本研究旨在探究粟酒裂殖酵母中ppr基因缺失引起细胞絮凝的机制,为研究粟酒裂殖酵母细胞絮凝的机制提供参考。
1 材料与方法
1.1 材料
1.1.1 菌株与质粒 粟酒裂殖酵母单倍体菌株yHL6381、Δppr3、Δppr4、Δppr6、Δppr10、Δppr3Δgsf2、Δppr4Δgsf2、Δppr6Δgsf2、Δppr10Δgsf2,pFA6a-hphMX6为本实验室保存。
1.1.2 培养基 YES培养基(每100 mL):葡萄糖3 g,酵母粉0.5 g,亮氨酸(L-leucine)20 mg,尿嘧啶(uracil)20 mg,组氨酸(histidine)20 mg,固体培养基加1.5~2.0 g的琼脂粉。
1.1.3 仪器与试剂 qRT-PCR试剂盒SYBR®Select Master Mix购自Takara(大连宝生物工程有限公司);反转录试剂盒iScriptTMcDNA Synthesis Kit购自BIO-RAD公司;Yeast Nitrogen Base Without Amino Acids购自Sigma公司;酵母提取物和蛋白胨购自Oxiod公司;其他化学药品除特殊说明外,均购自南京丁贝。
1.2 方法
1.2.1 絮凝分析 ①絮凝现象的显微观察[11]:将WT、Δppr3、Δppr4、Δppr6和Δppr10接种于YES液体培养基中,30 ℃摇床培养至稳定期,取适量的菌液制成装片,利用Zeiss Axio imager A1 microscope(Zeiss,Jena,Germany)(×63)观察细胞的分布情况。在剩余的菌液中添加EDTA至终浓度为0.01 mol/L,再取适量处理过的菌液制成装片用于显微镜观察。最后,在剩余的EDTA处理后的菌液中添加不同的金属离子至终浓度为0.2 mol/L,取菌液制成装片用于显微镜观察。②不同pH值条件下细胞絮凝程度的测定[20]:收集稳定期的WT、Δppr3、Δppr4、Δppr6和Δppr10细胞用ddH2O洗一遍;接着用6 mL 0.1 mol/L EDTA Na2重悬细胞10 min;再用ddH2O洗一遍。最后用各种pH值的琥珀酸Ca2+缓冲液(0.05 mol/L琥珀酸,0.05 mol/L Tris,0.008 mol/L CaCl2,分别用HCl 和NaOH调节至各个pH值)处理细胞10 min。震荡混匀,各取2 mL菌液转移至2个2 mL离心管中,分别设为对照组和实验组,小心吸取对照组上层液面200 μL细胞测OD600,记作At=0。实验组细胞静置10 min。小心吸取实验组上层液面200 μL细胞测OD600,记作At=10。计算细胞絮凝率(沉降率):V=1-At=10/At=0。
1.2.2 实时定量PCR 使用E.Z.N.A.®Yeast RNA Kit (OMEGA BIO-TEK) 抽提WT、Δppr3、Δppr4、Δppr6和Δppr10菌株的总RNA,用RNase-free DNase 消化提取样品中的DNA,用RevertAidTMFirst Strand cDNA Sythesis Kit (Fermentas) 进行反转录获得cDNA,将cDNA稀释一定倍数作为实时荧光定量PCR的模板,本研究以Actin作为内参,采用 ΔΔCT 法测定目标基因的相对表达量。计算公式:目标基因的相对表达量=2-ΔΔCT。
2 结果与分析
2.1 ppr3、ppr4、ppr6和ppr10敲除菌产生的细胞凝集现象
已知ppr3、ppr4、ppr6和ppr10的缺失能引起线粒体功能异常。在培养这四个缺陷菌的过程中发现单独敲除ppr3、ppr4、ppr6和ppr10基因会引起缺陷菌出现细胞凝集现象。因此,用蔡司显微镜对这些菌株的菌悬液进行观察,结果如图1所示,野生型yHL6381的细胞均匀分布在显微镜视野范围内,而Δppr3、Δppr4、Δppr6和Δppr10呈现出不同程度的细胞聚集现象。

图1 ppr3、ppr4、ppr6、ppr10的敲除引起细胞絮凝Fig.1 Deletion ofppr3,ppr4,ppr6andppr10cause aggregation of the mutant cells
2.2 ppr3、ppr4、ppr6和ppr10的敲除引起的细胞絮凝
无性絮凝的机制表明细胞表面的半乳糖残基可能是一种介导细胞之间发生凝集的受体,一旦培养基中存在游离的半乳糖就会对絮凝现象产生竞争性抑制。为了鉴定Δppr3、Δppr4、Δppr6和Δppr10敲除菌引起的细胞凝集是否属于无性絮凝,用含有半乳糖的培养基分别培养野生型菌株和这四个缺陷菌菌株。实验现象如图2A所示,在添加半乳糖的培养基中Δppr3、Δppr4、Δppr6和Δppr10细胞凝集现象消失,说明半乳糖可以抑制由这些敲除菌引起的细胞凝集。
为了验证本研究中的絮凝是否受Ca2+诱导,首先用EDTA处理突变体细胞,使其解絮凝,然后再添加CaCl2。结果如图2B所示, 添加EDTA可以使这些敲除菌解絮凝,但添加CaCl2后,絮凝又会再次形成,说明这种絮凝是可逆的并且依赖Ca2+。上述结果表明敲除ppr3、ppr4、ppr6和ppr10基因可以引起由Ca2+依赖型半乳糖结合蛋白参与的细胞絮凝。
2.3 不同pH值对Δppr3、Δppr4、Δppr6和Δppr10细胞絮凝的影响
溶液中的pH水平对细胞的絮凝会产生一定的影响。pH的改变主要通过影响细胞表面絮凝因子的蛋白活性进而对细胞的絮凝产生影响。对WT和Δppr3、Δppr4、Δppr6及Δppr10在不同pH情况下的絮凝程度分别进行了检测,如图3所示,这几株ppr缺陷菌在pH 3~pH 8之间絮凝程度最严重,而pH小于2时会极大地抑制细胞的絮凝。这一结果说明裂殖酵母细胞絮凝发生在接近生理条件的pH情况下。

图2 半乳糖和CaCl2对细胞的处理Fig.2 Cells were treated with CaCl2and galactose

图3 不同pH值对WT和Δppr3、 Δppr4、Δppr6、Δppr10絮凝的影响Fig.3 The effects of different pH to the flocclation of WT,Δppr3,Δppr4,Δppr6andΔppr10
2.4 不同金属离子对Δppr3、Δppr4、Δppr6和Δppr10细胞絮凝的影响
除了溶液中的pH水平,金属离子对于细胞的絮凝也会产生一定的影响。Ca2+是引发絮凝的一个关键因素,它可以通过维系絮凝因子最适的构像而可逆地激活絮凝因子的活性。同时絮凝现象的出现最终还要依赖暴露于细胞表面的糖蛋白与相邻细胞的半乳糖残基结合,进而形成细胞团沉淀到培养基中的底部。而Ca2+则直接参与了这种结合,它像个桥梁一样将糖蛋白与半乳糖残基紧紧连在一起[21]。那么其他的金属离子对于Δppr3、Δppr4、Δppr6和Δppr10的絮凝是否也能起到相同的作用?因此,设计实验来验证以上假设。用EDTA处理野生型yHL6381和Δppr3、Δppr4、Δppr6以及Δppr10,取出适量细胞液制成装片,作为自身空白对照。将剩余的细胞液用终浓度为0.2 mol/L阳离子(Ca2+、Mg2+、Na+、K+、Mn2+、Fe3+)处理后,取相同量处理后的细胞悬液制成装片,用显微镜观察。结果如图4所示,添加Ca2+、Mg2+、Na+或K+可以缓解这些突变体引起的絮凝,而Mn2+或Fe3+不可以。说明这种絮凝需要Ca2+、Mg2+、Na+或K+的参与,这些离子可能会激活絮凝因子。对照组,添加Fe3+可以诱导突变体和野生型都产生絮凝。
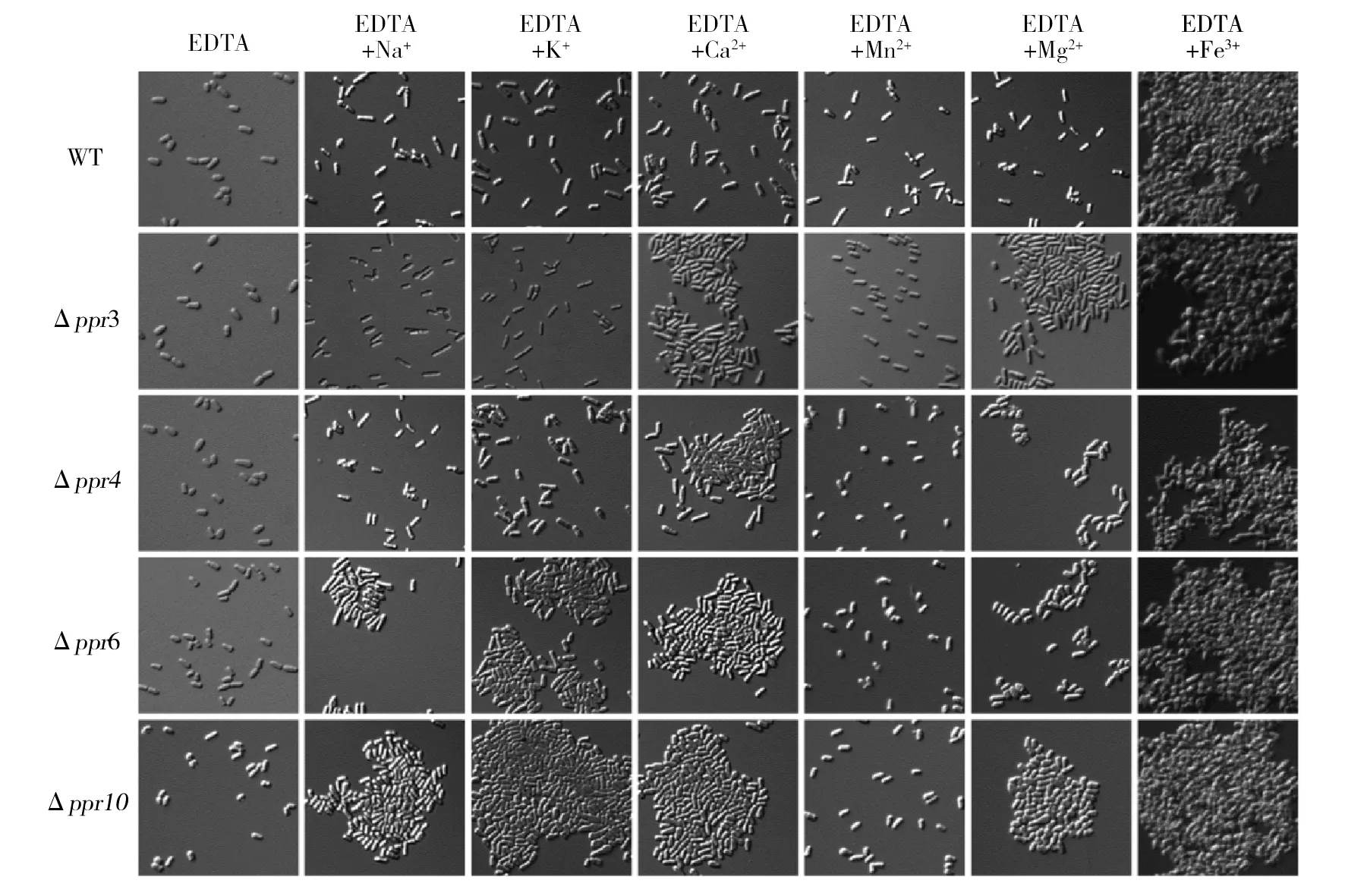
图4 不同金属离子对WT和Δppr3、Δppr4、Δppr6及Δppr10絮凝的影响Fig.4 The efects of different metal ions to the flocculation on WT,Δppr3,Δppr4,Δppr6 andΔppr10
2.5 ppr3、ppr4、ppr6和ppr10的敲除引起的编码絮凝因子基因表达上调
已经通过分析ppr10敲除菌的RNA-seq数据发现敲除菌中的基因表达发生改变。RNA-seq数据表明在Δppr10中参与絮凝的5个基因(gsf2、pfl6、pfl8、pfl9和cbf12)上调超过2倍[19]。其中pfl9和gsf2上调非常显著。为了验证这一结果,使用qRT-PCR来分析在ppr3、ppr4、ppr6和ppr10细胞中已报导的絮凝基因(gsf2、pfl2、pfl3、pfl4、pfl6、pfl7、pfl8、pfl9和SPBPJ4664.02)[10]。结果如图5A所示,gsf2、pfl6、pfl8和pfl9在突变体中表达上调,其中pfl9和gsf2上调的最为明显。因此,可以推断Δppr3、Δppr4、Δppr6及Δppr10是通过高表达细胞表面絮凝因子来实现絮凝。Gsf2是一个半乳糖特异性絮凝因子,在粟酒裂殖酵母中对无性絮凝起重要作用[10]。为了验证gsf2是否影响ppr敲除菌引起的絮凝,构建了双敲菌,在这些敲除菌的基础上敲除gsf2。结果发现敲除gsf2能够消除由ppr敲除菌引起的絮凝(图5B)。

图5 突变体菌株细胞中的一些絮凝基因的表达Fig.5 mRNA levels of some putative adhesion gene in wild-type,Δppr3,Δppr4,Δppr6and Δppr10 cells
2.6 ppr3、ppr4、ppr6和ppr10的敲除引起的编码细胞壁重构酶基因表达上调
在裂殖酵母和芽殖酵母中参与细胞壁重构酶的基因对絮凝起着重要作用[10,22]。用qRT-PCR检测ppr敲除菌中已知的4个编码细胞壁重构酶的基因agn2、psu1、gas2和Spac4h3.03c的表达。结果如图6所示,发现gas2和Spac4h3.03c表达上调明显高于agn2和psu1。
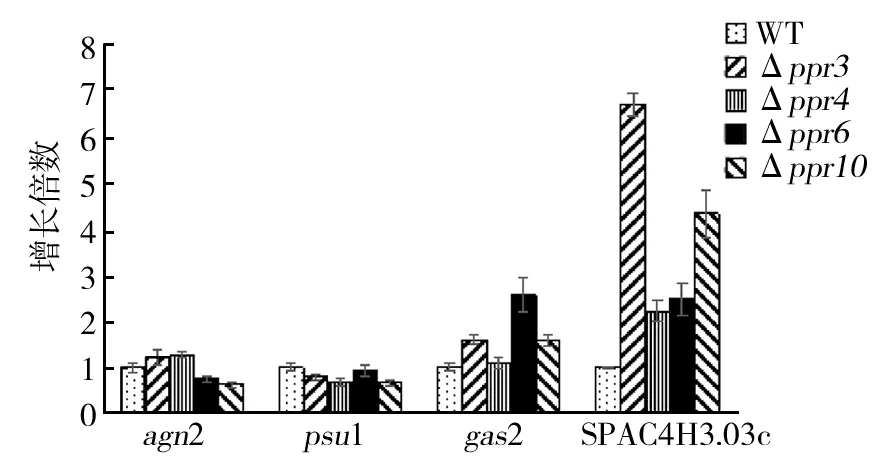
图6 突变体菌株细胞中编码 细胞壁重构酶基因的表达Fig.6 mRNA levels of cell wall remodeling enzyme gene in wild-type,Δppr3,Δppr4,Δppr6andΔppr10 cells
3 讨 论
粟酒裂殖酵母基因组编码10个PPR蛋白,这10个蛋白没有表现出序列上的相似性。除了Ppr5,失去其他几个蛋白中的任何一个,都会对线粒体DNA基因组的表达起负调控,引起线粒体ETC途径受损和呼吸缺陷。在ppr敲除菌中,只有Δppr3、Δppr4、Δppr6和Δppr10会引起无性絮凝表型。已有研究报道这四个ppr敲除菌在非发酵型培养基上表现出严重的生长缺陷,并且会引发线粒体功能异常从而导致体内铁离子失衡,进而促进体内ROS水平升高诱发细胞调亡[29]。因为絮凝是一种紧密的结构,细胞间存在着很小甚至没有间隙,所以线粒体功能严重受损会使细胞产生如絮凝这样的防御机制。因此,认为细胞絮凝是一种共保护机制来帮助细胞抵御环境压迫[23]。
在芽殖酵母中絮凝主要受到黏着蛋白Flo1、Flo5、Flo9、Flo10、Flo11、Fig2和Aga1的调控,这些基因的过表达会引起细胞絮凝,其中FLO1最为显著[24-25],但FLO1的表达受转录因子Flo8和Mss11的调节。在粟酒裂殖酵母中絮凝是由絮凝因子和细胞壁重构酶基因共同调节。本研究发现ppr3、ppr4、ppr6和ppr10敲除菌中絮凝因子表达明显上调,特别是gsf2和pfl9;细胞壁重构酶基因gas2和Spac4h3.03c的表达也明显上调。既然絮凝是由絮凝因子及细胞壁重构酶基因控制的,那么ppr3、ppr4、ppr6和ppr10的缺失可能会影响粟酒裂殖酵母的细胞壁特性。另外,很有可能在Δppr3、Δppr4、Δppr6和Δppr10中还有其他尚未确定的转录因子也参与了絮凝基因的上调。还需实验进一步探究gas2和Spac4h3.03c的表达上调是如何进行的。
本研究发现ppr3、ppr4、ppr6和ppr10的缺失会引起由Ca2+依赖型半乳糖结合蛋白参与的细胞絮凝。因此,对ppr基因敲除引起的细胞絮凝机制产生了兴趣。研究发现pH值和金属离子是影响这类缺陷菌细胞絮凝的因素。通过qRT-PCR技术得出,ppr3、ppr4、ppr6和ppr10的缺失会使细胞内编码絮凝因子和细胞壁重构酶基因表达上调。综上所述,线粒体功能异常会诱导絮凝基因表达上调和细胞壁属性发生改变,从而产生絮凝现象。
