不同坡位对4种阔叶乡土树种叶片养分的影响
2019-05-13王卓敏
余 明,王卓敏,薛 立
(华南农业大学 林学与风景园林学院,广东 广州 510642)
叶片能灵敏反映植物对环境变化的适应性和生长发育状况[1],是植物储存养分和发挥同化作用的重要器官,其养分含量高低可以间接反映林分和土壤的养分水平,是对森林生态系统进行养分研究的重要依据[2]。植物叶片养分受到如气候条件、生长阶段、季节动态和微环境因子等诸多因素的影响[3-5],坡位作为较小尺度上的地形因子,通过影响土壤、光照、降水等环境因素而影响着植物的生长和养分特点[6]。另外,不同树种的叶片结构和功能存在着差异,导致对养分的需求不同[7]。
前人关于叶片养分的研究多集中于叶片养分特征[2]、叶片养分的空间分布格局[8]和动态变化[4]、叶片养分与土壤养分的关系[9-10]、叶片养分诊断[11]、施肥对叶片养分的影响[12-13]等方面,如冯梅等[14]报道了胡杨叶形变化与叶片养分间的关系,刘宏伟等[15]发现了重庆石灰岩地区主要木本植物叶片的养分再吸收规律,王卓敏等[16]研究了广东横琴岛园林植物叶片的养分特性,方怡然等[17]研究了广东云勇林场3 种人工阔叶林的叶片养分状况,刘落鱼等[18]对10 种茶花的叶片养分特征进行了探索,戚嘉敏等[19]研究了油茶叶片氮磷钾养分的年动态变化规律。但鲜见不同坡位的叶片养分变化的报道[6],而有关华南地区坡位对于叶片养分的研究尚未有报道。木荷Schima superba、灰木莲Manglietia glauca、乐昌含笑Michelia chapensis、山杜英Elaeocarpus sylvestris是华南地区主要乡土树种,通过研究它们在不同坡位上的叶片养分差异,可以揭示这些树种在不同坡位的适应特征,为改善华南地区的生态环境提供科学依据。
1 材料和方法
1.1 试验地概况
研究地点位于韶关市小坑国家森林公园。试验地土壤类型为砂页岩发育成的红壤,土层较深厚,海拔为200 ~300 m,属于亚热带季风气候,年均温度20.1 ℃,最热月份平均气温28.9 ℃,极端最高气温39.5 ℃,最冷月份平均气温9.6 ℃,极端最低气温-5.3 ℃,年活动积温7 300℃,年平均降水量1 640 mm。2016年4月栽植木荷、灰木莲、乐昌含笑、山杜英幼苗44 hm2(每个树种11 hm2),栽种密度为900 株·hm-2(2 m×3 m)。2016年在试验地沿坡面分别在上坡、中坡、下坡各设置面积为20 m×20 m 的3 个样地。各样地幼苗规格见表1。

表1 4种阔叶树种幼苗的基本情况Table1 General status of seedlings of four broadleaf tree species
1.2 试验方法
2018年的3月份采集幼树叶片带回实验室进行叶片碳、氮、磷的测定。在各样地内采摘4 个树种完整且有代表性的成熟叶30 枚,放入密封袋保鲜并拿回实验室。将叶片放入烘箱,105 ℃杀青20 min 后在85 ℃烘干到恒质量,并测干质量。用重铬酸钾容量法测定碳含量,半微量凯氏法测定氮含量,铝蓝比色法测定磷含量[20-21]。
1.3 数据处理
植物叶片C、N、P 含量均采用质量含量(g·kg-1) ,C ∶N、C ∶P、N ∶P 均为质量比,采用Microsoft Excel 2007 及SPSS 软件进行数据处理和图表绘制。
2 结果与分析
2.1 4 个树种在不同坡位的叶片养分含量对比
荷木的叶片碳含量(g·kg-1)为上坡(525.34)显著高于中坡(505.65)和下坡(507.01);灰木莲为上坡(527.36)和下坡(527.18)显著高于中坡(501.99);乐昌含笑为上坡和中坡显著低于下坡,其叶片碳含量在上坡、中坡、下坡分别为495.58、496.33、515.32 g·kg-1;山杜英为下坡(473.85)<上坡(483.58)<中坡(507.58),各坡位间差异显著 (P<0.05)(图1)。

图1 4 个树种不同坡位的叶片碳含量Fig.1 C content in leaves of four tree species at different slopes
荷木和山杜英的叶片氮含量(g.kg-1)均为中坡最低,下坡最高,在各坡位间差异显著,荷木的叶片氮含量(g·kg-1)为中坡(12.34)<上坡(14.66)<下坡(16.59),山杜英为中坡(12.82)< 上坡(14.86)<下坡(18.38);灰木莲和乐昌含笑均为中坡最高,上坡最低,各坡位间差异显著,上、中、下坡叶片氮含量分别为11.27、16.94、16.43 和10.31、12.64、11.37 g/kg(P<0.05)(图2)。
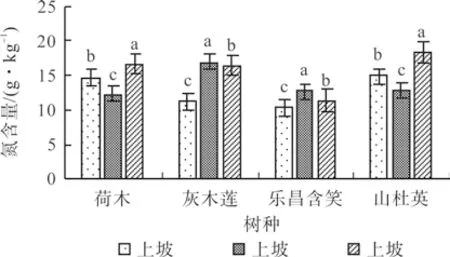
图2 4 个树种不同坡位的叶片氮含量Fig.2 N content in leaves of four tree species at different slopes
荷木的叶片磷含量(g·kg-1)为中坡(0.82)<下坡(0.99)<上坡(1.05),各坡位间差异显著(P<0.05);灰木莲和乐昌含笑均为上坡<下坡<中坡,且差异显著(P<0.05),其中灰木莲上、中、下坡的叶片磷含量分别为0.77、1.14、1.07 g·kg-1,乐昌含笑为0.65、0.98、0.93 g·kg-1;山杜英的叶片磷含量(g·kg-1)表现为中坡(0.85)<上坡(0.91)<下坡(1.44),各坡位间差异显著(P<0.05)(图3)。
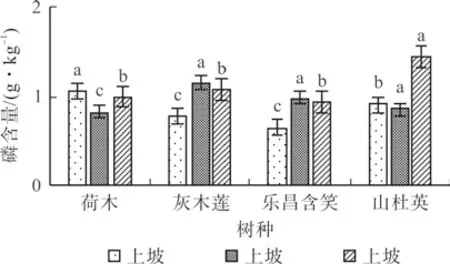
图3 4 个树种不同坡位的叶片磷含量Fig.3 P content in leaves of four tree species at different slopes
2.2 同一坡位4 个树种的叶片养分含量
在上坡,4 个树种的叶片碳含量(g·kg-1)顺序为灰木莲(527.36)>荷木(525.34)>乐昌含笑(495.58)>山杜英(483.58),除荷木和灰木莲之间差异不明显外,其余树种间均差异显著(P<0.05)(图4A)。
在中坡,叶片碳含量(g·kg-1)为山杜英最高(507.58),荷木(505.65)、灰木莲(501.99)、乐昌含笑(495.6)次之(图4B)。
在下坡,叶片碳含量(g·kg-1)按灰木莲(527.18)、乐昌含笑(515.32)、荷木(507.01)、山杜英(473.85)依次降低,树种间差异显著(P<0.05)(图4C)。

图4 4 个树种在同一坡位的叶片碳含量Fig.4 C content in leaves of four tree species at the same slope
上坡的叶片氮含量(g·kg-1)顺序为山杜英(14.86)>荷木(14.66)>灰木莲(11.27)>乐昌含笑(10.31),其中荷木和山杜英的叶片氮含量显著高于其余树种(P<0.05)(图5A)。中坡的叶片氮含量为灰木莲最高(16.94),山杜英(12.82)与乐昌含笑(12.64)次之,荷木最低(12.34)(图5B)。下坡的叶片氮含量按照山杜英(18.38)、荷木(16.59)、灰木莲(16.43)、乐昌含笑(11.37)的顺序依次降低,除荷木和灰木莲之间外,其余树种间差异显著(P<0.05)(图5C)。
上坡的叶片磷含量(g·kg-1)为荷木最高,乐昌含笑显著低于其余树种,顺序为荷木(1.05)>山杜英(0.91)>灰木莲(0.77)>乐昌含笑(0.65) (P<0.05)(图6A);中坡的叶片磷含量按灰木莲(1.14)、乐昌含笑(0.98)、山杜英(0.85)、荷木(0.82)依次降低(图6B);下坡的叶片磷含量为山杜英(1.44)>灰木莲(1.07)>荷木(0.99)>乐昌含笑(0.93),4 个树种间有显著差异(P<0.05)(图6C)。
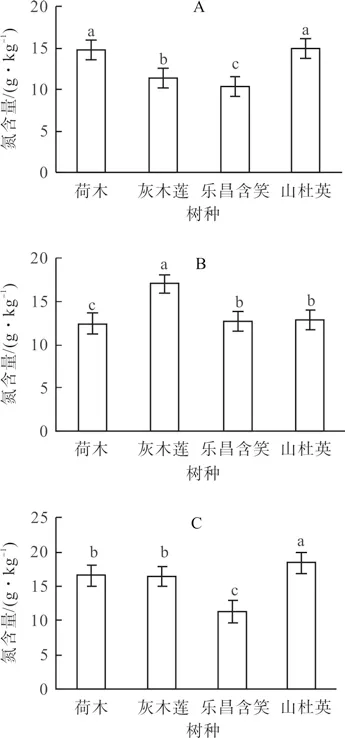
图5 4 个树种在同一坡位的叶片氮含量Fig.5 N content in leaves of four tree species at the same slope
2.3 植物叶片C、N、P 特征

图6 4 个树种在同一坡位的叶片磷含量Fig.6 P content in leaves of four tree species at the same slope
各树种在不同坡位的植物叶片C、N、P 的化学计量比如表2所示。4 种植物叶片C ∶N 在上坡、中坡、下坡的分布范围分别为32.5 ~48.1、29.6 ~41.0、25.8 ~45.3,均高于全球平均水平(22.5);C ∶P 在各坡的分布范围分别为500.1 ~766.0、439.9 ~614.0、329.6 ~557.0;N ∶P 在各坡位的分布范围分别为14.0 ~16.4、12.9 ~15.1、12.3 ~16.8,其中中坡与下坡的乐昌含笑以及下坡的山杜英均低于中国树种的叶片平均氮磷比(14.4)[22]和全球平均水平(13.8)[23]。

表2 4个树种在各坡位的叶片化学计量比Table2 Ratios of C,N and P contents in four different tree species leaves at different slope positions
3 结论与讨论
3.1 不同坡位的叶片养分特征
本研究中,灰木莲和乐昌含笑的叶片N、P 含量均为中坡最高,山杜英的叶片N 和P 含量均为下坡最高,荷木则在中坡最差。各坡位的植物叶片N 含量均低于全国平均水平(20.2 mg·g-1)以及全球水平(20.1 mg·g-1), 除了下坡生长的山杜英外,其余树种的叶片P 含量均未达到全 国平均水平(1.21 mg·g-1)及全球水 平(1.42 mg·g-1)[23]。各树种的叶片N 和P 含量整体水平较低,这可能是因为华南地区降雨丰富导致移动性强的有效态氮容易发生淋溶而流失所致[24]。姜沛沛等[25]的研究表明,植物体内养分含量体现了植物对环境的适应特征[26],植物体中N、P 元素主要来源于土壤,叶片N 和P 含量也与土壤中养分含量密切相关[27]。华南地区高温多雨的气候使土壤风化强烈,引起土壤氮和磷的缺乏,可能导致叶片N 和P 含量较低。
坡位对4 种树种的土壤养分通常有显著影响。坡位通过影响光照、温度、降水等环境因子的分布而影响植物的生长[6],也通过水肥的再分配而影响林分土壤养分。侵蚀过程将上坡的表层土壤细粒搬运到下坡后沉积, 造成上坡土壤贫瘠,下坡土壤湿润肥沃。因此,不同坡位的植物特征均有较大差异[28]。本研究中,N 和P 含量通常在上坡较低,中坡和下坡较高,与土壤肥力的状况相同。灰木莲和乐昌含笑的叶片N、P 含量均为中坡最高,可能 是由于下坡的林下植物生长茂盛,竞争力较强,限制幼苗的光合营养空间, 其密集的根系阻碍了幼树根系的伸展,减少了其对养分和水分的吸收[29]。
在上坡,荷木的叶片N 和叶P 含量均较高;在中坡,灰木莲叶片N 和P 含量最高;在下坡,山杜英叶片N 和P 含量为最高,乐昌含笑最低。在植物生长过程中,叶片的养分含量变化最为敏感,其浓度的变化由许多因素共同决定,如遗传因素、物候期和外界环境等[4]。在本研究中,不同树种的叶片N 和P 存在差异,可能是因为不同树种对养分的需求不同所致,因为不同树种的遗传特性导致对N、P 需求和选择吸收上的差异[7]。同一坡位的一些树种的叶片N、P 含量无显著差异,原因可能是叶片养分含量受土壤养分、光照、温度和水分的复杂影响所致。
3.2 树木叶片的化学计量特征
植物叶片C、N、P 等化学元素计量特征能够充分反映植物在新陈代谢和生长发育过程中对营养成分的利用情况以及对贫瘠环境的适应能力[30],其变化特征与生长率有很大的关系[31-32],可以反映植物生长过程中对于资源的利用及土壤养分循环的状况,揭示植物与环境之间相互平衡制约的关系及其生长是否受到相关养分的限制[33]。其中叶片C ∶N 和C ∶P 代表着植物固碳能力的高低[34],反映了植物对营养元素的利用效率和植物生长速度[35]。本研究中,4 种树种叶片在各坡位C:N 值范围为29.6 ~51.5,均高于全球平均水平(22.5)。C ∶P 比的范围在329.6 ~766.0之间,除上坡的灰木莲和乐昌含笑外,均低于我国浙江天童山常绿阔叶林(758.0) 和常绿针叶林(677.9)[36]。俞月凤等[37]研究发现,拥有较高的C ∶N 和较低C ∶P 的植物在单位N、P 养分条件下,其叶片的N 利用效率高,而P 利用效率低。N、P 是细胞结构与功能最为重要的生命元素,植物叶片的N ∶P 被认为可以作为判断植物生长氮受限或磷受限的指标[38],反映了植物特定的生长策略[39]。N ∶P 值小于14 通常意味着氮受限,而N ∶P 值大于16 则意味着植物更多的受到磷的限制,介于两者中间表明受到氮、磷元素的共同限制作用。本研究中,上坡的山杜英以及下坡的荷木的N ∶P 比均大于16,可见它们的生长受到磷限制,而中坡与下坡的乐昌含笑以及下坡的山杜英均低于14,意味着受到氮的限制,其余各坡位植物则受到氮和磷的共同限制[40],因此可以适当追加N、P 肥,通过人为干预促进幼树的生长,以便发挥良好的生态效益[41]。
