肠外致病性大肠杆菌LuxR家族转录调节蛋白YjjQ研究进展
2019-05-10傅丹丹肖亚婷祁克宗宋祥军
傅丹丹,肖亚婷,祁克宗,宋祥军,涂 健,薛 媚
(安徽农业大学动物科技学院/兽医病理生物学与疫病防控安徽省重点实验室,安徽 合肥 230036)
致病性大肠杆菌包括肠道内致病性大肠杆菌及肠外致病性大肠杆菌,肠外致病性大肠杆菌包括:尿道致病性大肠杆菌(UropathogenicEscherichia coli,UPEC)、新生儿脑膜炎大肠杆菌(NeonatalmeningitisEscherichia coli,NMEC)、禽致病性大肠杆菌(Avian PathogenicEscherichia coli,APEC)等[1]。肠外致病性大肠杆菌常引发腹泻、尿路感染、新生儿脑膜炎和败血症等多种疾病,给人和动物的健康和公共卫生带来严重威胁[2]。但由于APEC及UPEC等肠外致病性大肠杆菌血清型众多,且毒力因子复杂,导致其致病机制尚不明确。
转录因子Y jjQ是LuxR家族的一种调控蛋白,其含有 LuxR家族共有的螺旋 -转角 -螺旋(helix-turn-helix,HTH)DNA结合结构域,可通过结合靶基因的启动子序列调控相关基因的转录[3]。LuxR型转录调节因子(LuxR-type transcriptional regulators)广泛存在于革兰氏阴性菌中,包含YjjQ、BglJ、RcsA-RcsB等[4-5],在细菌群体感应、生物被膜形成及胞外酶、毒力因子和次生代谢产物的合成中发挥重要作用。yjjQ常与bglJ构成操纵子以发挥转录调控作用,该操纵子可被全局阻遏蛋白H-NS抑制,而被转录激活因子 LeuO激活[3](图 1),H-NS和 LeuO对 yjjQ-bglJ操纵子的调控表明这些基因与应激反应和/或毒力相关性状的控制有关[6]。本文依据已有的研究详述了肠外致病性大肠杆菌LuxR家族转录调节蛋白Y jjQ的研究进展,为探索相关转录因子对致病菌致病调控机制提供理论支撑。

图1 LuxR家族转录调控网络[7]Fig.1 Transcriptional regulatory network of the LuxR family[7]
1 yjjQ的位置
yjjQ是yjjQ-bglJ操纵子的第一个基因[6],其3'末端与下游基因bglJ 5'末端有42 bp开放阅读框串联重叠,其5'末端与膜蛋白基因yjjP相邻,且yjjP与yjjQ之间存在一段含有yjjQ启动子和调节元件的557 bp的非编码区[8-10](图2)。目前对Y jjQ的蛋白构象和功能域还处于未知阶段,因此进一步探究其功能域有助于助揭示其在致病机制中所发挥的作用。
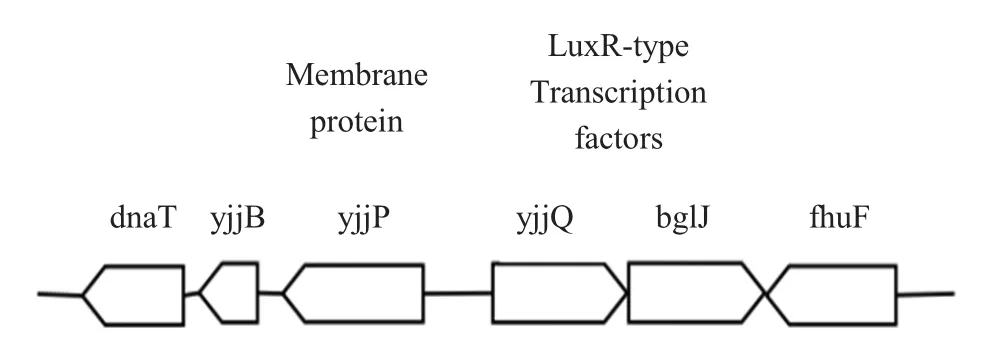
图2 yjjQ-bglJ操纵子[3]Fig.2 yjjQ-bglJoperon[3]
2 YjjQ对靶基因的转录调控作用
研究表明,转录因子YjjQ是转录调控网络的重要调节剂,其通过结合靶基因启动子序列而抑制靶基因转录,主要对包括鞭毛形成、生物被膜形成能力、摄铁系统、胞外酶和毒力因子等相关基因的调控。
2.1 抑制细菌鞭毛及生物被膜的形成 鞭毛(Flagellum)作为细菌最主要的运动器官,参与细菌的黏附侵袭、生物被膜形成、分泌毒力因子和细菌运动等[11]。研究表明,鞭毛的合成和功能的发挥至少涉及50多个基因,这些基因共形成3个调控等级,分布在10多个操纵子上。其中flhDC操纵子位于鞭毛调控系统的最高等级,其主要编码鞭毛主调节子FlhD4C2,在鞭毛合成、细菌分裂和毒力等方面发挥核心作用[5,12]。yjjQ过表达可导致大肠杆菌K-12、APEC IMT5155菌的运动能力显著降低,且研究者通过构建LacZ报告基因融合体发现其10倍抑制了大肠杆菌K-12flhDC基因的转录活性[6,8],这表明yjjQ对鞭毛形成有显著影响。
生物被膜(Biofilm,BF)主要由细菌及其分泌的黏附蛋白、胞外多糖等组分构成,细菌利用生物被膜可增强自身耐药性和抵抗外界环境压力的能力。据统计,超过65%的微生物感染与生物被膜有关[13]。研究发现,yjjQ抑制大肠杆菌K-12和UPEC 536生物被膜的形成,且通过调控膜相关基因的转录参与生物被膜、荚膜及其它膜结构的功能和稳定性,进而影响细菌毒力[6,10]。
2.2 抑制铁摄取基因的表达 铁元素是大部分细菌生存所必需的微量元素,参与体内的多种酶催化反应,对机体的生命活动起重要作用,但铁过量时对细菌有毒害作用[14]。在长期进化中,为维持体内游离铁的浓度,细菌形成一系列螯合和摄取铁元素的系统,以保证其各种功能的正常发挥。研究表明,摄铁系统与细菌毒力和生物被膜形成有密切的联系[8,15]。当APEC IMT5155菌株缺失yjjQ基因后,发现摄铁相关基因ireA,fyuA,sitA和chuA的表达均下调[8],其中chuA参与病原菌血红素摄取;周质铁结合蛋白SitA介导锰和铁运输,与细菌毒力相关[16-17]。
2.3 转录调控其它靶基因的表达 yjjQ还参与调控其它多种基因的表达:①YjjQ抑制氨基酸合成酶基因,如与芳香族氨基酸合成有关的tyrB和aroA基因,参与催化α,β-羟基酸的脱水反应的ilvD基因(编码二羟基酸脱水酶)[18],与琥珀酰转移酶相关的dapD基因等[19]。Li等人在敲除APEC IMT5155菌株的yjjQ基因后发现上述的氨基酸合成酶基因活性均下降,并在M 9培养基中加入0.4%酪蛋白氨基酸后缺失株的生长能力恢复至野株型水平[8],表明YjjQ参与氨基酸代谢并调控细菌的生命代谢;②调控膜蛋白基因,如ompC、ompT、gfc、dcsA、tolC、kpsMT等。外膜蛋白TolC与细菌耐药、毒力、耐酸性有关,常作为药物靶标进行研究[20];OmpC是大肠杆菌重要的外膜蛋白,参与细胞外膜形成,可增强机体的免疫力和耐药性[21],这些膜蛋白通常位于细菌细胞壁的外膜(OM)和/或内膜(IM),表明yjjQ影响细菌生物膜结构及细胞壁完整[6,8,10];③转录调控其它基因,如与脂多糖LPS相关的基因rfaE,与耐酸性相关基因adiY,与能量代谢相关基因cra,与耐酸性和致病性相关基因dctR,与锌吸收有关的zur基因[8,10,22]。
目前已发现Y jjQ调控的基因(部分靶基因如表1)在病原菌转录调控、新陈代谢等方面发挥重要作用,但主要集中于对鞭毛合成抑制作用的研究,对其它致病因子及其在整个调控网络中扮演的角色仍有待深入探究。

表1 YjjQ转录调控的部分靶基因及其主要功能Table 1 Transcriptional Regulation of Some Target Genes and Their Major Functions
3 YjjQ的转录调控机制
病原菌感染宿主细胞涉及到多种毒力因子的共同作用,致病过程复杂,而转录因子(Transcription factor,TF)是一类能够调控细菌生命活动的蛋白质[26],不仅可以调控下游靶基因,也受其自身的上游调控子的调控,在细菌复杂有序的级联网络结构中处于重要位置。YjjQ对靶基因转录抑制主要有两种机制:①通过YjjQ的HTH DNA结合结构域直接与靶基因启动子的序列结合,进而干扰靶基因与转录因子,或与RNA聚合酶结合以抑制转录发生;②通过抑制其它转录调节因子的表达而间接起作用,如假定的靶基因ucpA和fruBKA。cra分别是ucpA转录激活子和fruBKA转录抑制子,Y jjQ通过抑制cra可以间接下调ucpA的表达及上调fruBKA表达[27-28]。
4 展望
肠外致病性大肠杆菌APEC和UPEC均能够引起严重的肠外疾病,且有研究提示APEC和UPEC可能互为毒力基因库,但目前关于APEC及UPEC致病机制尚不明确。YjjQ通过抑制与菌毛合成、鞭毛合成和生物被膜形成的相关基因,不仅影响APEC和UPEC对宿主细胞的入侵和粘附,甚至在尿道感染的复发中起重要作用。
群体感应(Quorum Sensing,QS)和二元调控系统(Two Component System,TCS)与病原菌的致病性和耐药性等密切相关,但目前已有的研究大多针对单个毒力基因或调控系统。LuxR类转录因子不仅是群体感应过程中一个重要参与者,且结构上与二元调控系统的调控蛋白有一定的相似性。Y jjQ是否可与群体感应及二元调控系统等在病原体网络调控中协同发挥作用,以调节细菌行为从而适应周围环境,尚未见报道。将转录因子、群体感应系统和二元调控系统等综合考虑,不仅将为探究禽致病性大肠杆菌、尿道致病性大肠杆菌等肠外致病性大肠杆菌致病机制提供新思路,同时为探究禽致病性大肠杆菌与尿道致病性大肠杆菌是否互为对方毒力基因库提供理论依据,同时为研究其它病原菌致病机制及相互间的关系提供线索。
