中间偃麦草第6同源群特异STS标记开发
2019-04-23刘淑娟张晓军李欣刘成白建荣任永康郑军李世姣郭慧娟梅超张树伟畅志坚乔麟轶
刘淑娟,张晓军,李欣,刘成,白建荣,任永康,郑军,李世姣,郭慧娟,梅超,张树伟,畅志坚*,乔麟轶*
(1.山西大学生物工程学院,山西 太原 030006;2.山西省农业科学院作物科学研究所作物遗传与分子改良山西省重点实验室,山西太原 030031;3.山东省农业科学院作物研究所,山东 济南 250100; 4.山西省农业科学院小麦研究所,山西 临汾 041000)
中间偃麦草(Thinopyrumintermedium,2n=6x=42,JJJsJsStSt)是禾本科小麦族偃麦草属的多年生异花授粉植物,具有长势茂盛、适应性广、茎叶富含蛋白等特性,在世界各地被作为优良牧草而广泛栽培,同时也是防风固沙、保持水土和改良盐碱地的理想物种[1]。中间偃麦草属于部分异源-同源六倍体,其进化过程和染色体组成较为复杂。自1936年起,研究人员先后提出了 AXY[2]、BEF[3]、E1E2X[4-5]和B2X1X2[6]等染色体结构,目前普遍认为其组成应为JJsSt,其中 J 来自百萨偃麦草(Th.bessarabicum,2n=2x=14,JbJb)和二倍体长穗偃麦草(Th.elongatum,2n=2x=14,JeJe),St 来自拟鹅观草(Pseudoroegneiriastrigosa,2n=2x=14,StSt),Js则是与St发生部分重组的J染色体组[7-12]。
由于中间偃麦草与普通小麦(Triticumaestivum,2n=6x=42,AABBDD)同为六倍体,容易与之杂交成功,且因其抗逆性强,尤其对小麦白粉病、锈病、黄矮病、黑穗病等主要病害均表现出优良的抗性[1,13-22],因此成为小麦遗传改良的重要野生资源之一。目前国内外已将中间偃麦草诸多优良抗性导入小麦,并利用细胞学技术鉴定出一大批小麦-中间偃麦草附加系、代换系和易位系[14-22],其中,中间偃麦草第6同源群已被证实对小麦白粉病和条锈病免疫[16-17]。
随着分子生物学的发展,分子标记也被用于鉴定杂交材料中的外源染色体片段。Qi 等[21]利用1个 SSR 标记和3个 EST 标记鉴定出小麦抗白粉病材料 SN100109中附加了1对中间偃麦草2J 染色体;Salina等[17]利用9个 SSR 标记鉴定出小麦品系Tulaikovskaya的6D 染色体被同源的中间偃麦草6号染色体代换从而获得了抗病性;Li 等[22]利用42个 PLUG 标记鉴定出4个小麦-中间偃麦草附加系、代换系和易位系。然而由于中间偃麦草基因组尚未完成测序,而现有的中间偃麦草特异分子标记很大部分来自小麦等物种[15-22],或者是基于中间偃麦草某条染色体或染色体区段而开发的若干标记[23],远不能满足鉴定需要。为此,本研究对近期公布的中间偃麦草 GBS(genotyping-by-sequencing)芯片探针序列[24]进行组装和过滤,据此开发了一套分子标记,并对第6同源群的标记进行了筛选验证,以期为中间偃麦草染色体的鉴定提供较为经济和便捷的工具。
1 材料与方法
1.1 试验材料
中间偃麦草 Z1141、百萨偃麦草、拟鹅观草、普通小麦农家品种中国春、兰花麦、白秋麦、小白芒、江西早由作物遗传与分子改良山西省重点实验室提供;二倍体长穗偃麦草以及中国春-二倍体长穗偃麦草代换系6Je/6A、6Je/6B 和6Je/6D 由四川农业大学马建副研究员和山东省农业科学院刘成副研究员惠赠;小麦-偃麦草代换系材料 F881(6St/6D)由电子科技大学杨足君教授惠赠。试验时间为2018年3-10月。
1.2 序列组装及过滤
从NCBI数据库(https://www.ncbi.nlm.nih.gov/sra/)下载中间偃麦草 GBS 芯片探针数据(注册号SRX3008333),使用 SOAPdenovo 软件将其组装为 Contig 序列,并去除冗余,将所获序列与小麦中国春基因组(IWGSCv1.0,https://urgi.versailles.inra.fr/)进行比对,去除相似性大于80%的序列。
1.3 分子标记开发
对中间偃麦草Contig序列开发序列标签位点(sequence-tagged site,STS)分子标记,用于分析扩增产物的有无或长短。设计引物所用的 Perl-primer 脚本程序由山西省农业科学院王长彪副研究员惠赠,参数设置为:引物长度20 bp,产物长度范围100~400 bp。引物由生工生物工程(上海)股份有限公司合成。
1.4 DNA提取及标记验证
将试验材料在培养皿中发苗,待生长至3叶期剪取叶片。采用 CTAB 法提取材料叶片的基因组总 DNA。PCR 反应总体系为10 μL:1.5 μL DNA(50 ng·μL-1), 1.0 μL Primers(50 ng·μL-1), 5.0 μL Taq PCR Mix 预混液,2.5 μL ddH2O。反应扩增程序为:95 ℃ 5 min;95 ℃ 30 s、60 ℃ 30 s、72 ℃ 25 s,共35个循环;72 ℃ 10 min。扩增产物在8%非变性聚丙烯酰胺凝胶(Acr∶Bis=29∶1)中电泳后用0.1%硝酸银染色显影。
2 结果与分析
2.1 中间偃麦草序列组装及标记开发
利用下载的中间偃麦草 GBS 探针序列(每条60 bp)共组装出5877409条 Contig,长度范围为100~3094 bp,总碱基数915311073 bp(表1)。通过信息学分析,筛选出具有染色体同源群定位信息[24]的非冗余 Contig 序列5452条,在中间偃麦草第1至第7同源群分布数目依次为704、686、877、656、893、624、1012条(图1)。
对筛选出的5452条中间偃麦草 Contig 设计引物,共开发标记2019个,在第1至第7同源群分布数目依次为250、215、323、253、323、253和402个(图1)。
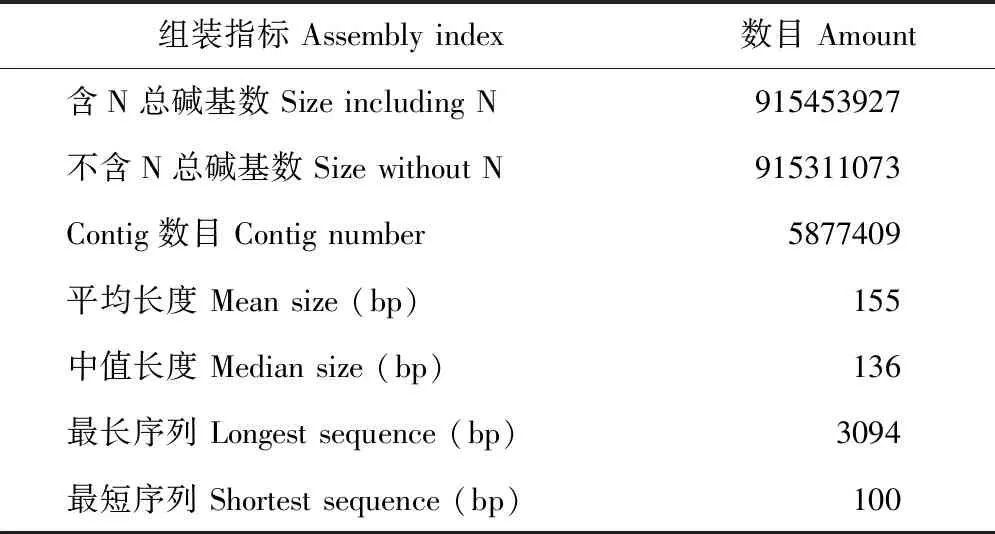
表1 中间偃麦草GBS探针序列组装结果Table 1 Sequence assembly for GBS probes of Th. intermedium
2.2 第6同源群标记筛选
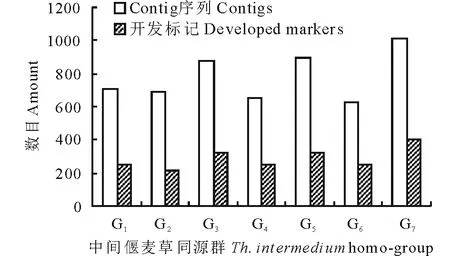
图1 中间偃麦草Contig序列和开发标记分布Fig.1 Distribution of Contigs and developed markers in Th. intermedium
利用5株中间偃麦草和5个小麦农家种的 DNA 对中间偃麦草第6同源群的253个分子标记进行筛选,共有160个标记扩增出差异条带(表2,图2),分布在同源群内的 Group6 (G6)-Chr1、G6-Chr2和G6-Chr3染色体,分别有58、59和43个,其中只在中间偃麦草中具有扩增产物的“+/-”型特异标记共有53个,依次为32(G6-Chr1)、13(G6-Chr2)和8(G6-Chr3)个。
2.3 G6特异标记所在基因组分析
利用中间偃麦草祖先种和6Je代换系对160个 G6特异标记进行检测,结果显示 G6-Chr2上59个标记中有11个标记在祖先种中无扩增或无相同条带,剩余标记在 Je基因组(二倍体长穗偃麦草、代换系 6Je/6A、6Je/6B 和6Je/6D)、Jb基因组(百萨偃麦草)和 St 基因组(拟鹅观草)有特异性扩增条带的数目依次为1、5和34,其中 St 特异标记占85%(图2b,图3),因此推断 G6-Chr2为中间偃麦草6St 染色体;而 G6-Chr1和 G6-Chr3上的 Je/Jb/St 基因组特异标记数目分别为10/1/18和6/4/11,暂不能确定其染色体组成。
2.4 Thi-6St特异标记用于材料验证
选择6St(G6-Chr2)染色体上的13个“+/-”型特异标记对代换系材料 F881(6St/6D)进行进一步验证。这些标记在中间偃麦草中都能扩增出条带,在 F881中能扩出11个(85%),在中国春中则无扩增(图4),该结果可从分子水平上验证代换系材料 F881中导入了6St 染色体,证实了Thi-6St 特异标记用于外源鉴定的可行性。
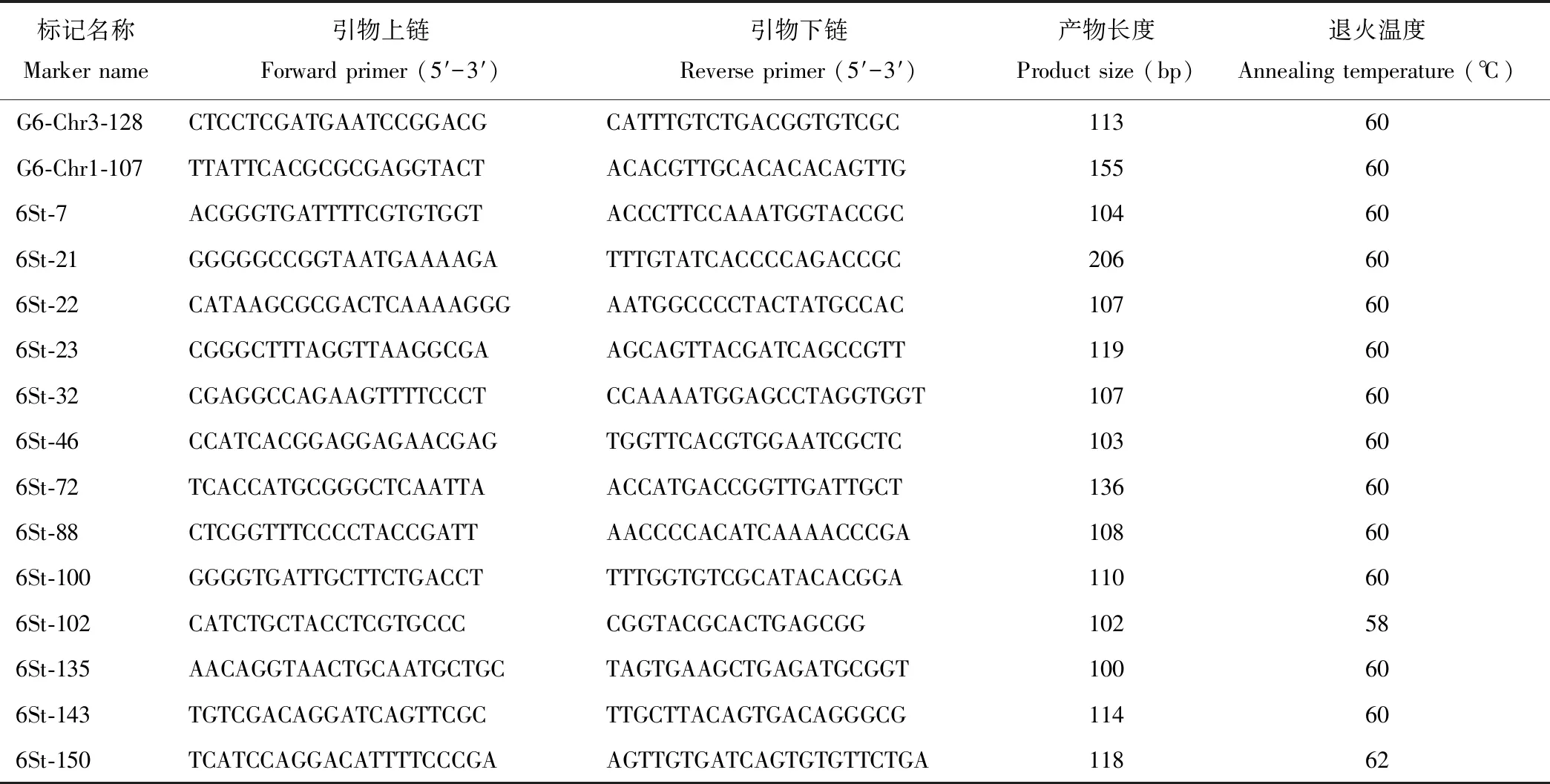
表2 中间偃麦草第6同源群部分特异分子标记Table 2 Parts of specific molecular markers in the sixth homo-group of Th. intermedium
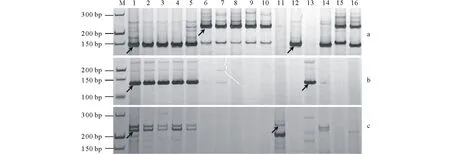
图2 第6同源群特异标记G6-Chr1-107(a)、G6-Chr2-72(b)、G6-Chr3-128(c)的鉴定结果Fig.2 Identification results of Thi-G6 specific markers G6-Chr1-107(a), G6-Chr2-72(b) and G6-Chr3-128(c) M: 指示带;1~5:中间偃麦草 Z1141-1~5;6~10:普通小麦农家种兰花麦、白秋麦、小白芒、江西早和中国春;11:百萨偃麦草;12:二倍体长穗偃麦草;13:拟鹅观草;14~16:中国春-二倍体长穗偃麦草代换系6Je/6A、6Je/6B和6Je/6D。箭头指示多态性条带。M: Marker ladder; 1-5: Th. intermedium Z1141-1~5; 6-10: Wheat landraces Lanhua, Baiqiu, Xiaobaimang, Jiangxizao and Chinese spring; 11: Th. bessarabicum; 12: Th. elongatum; 13: Ps. strigosa; 14-16: Chinese spring-Th. elongatum substitution line 6Je/6A, 6Je/6B and 6Je/6D. Arrows show polymorphic bands.
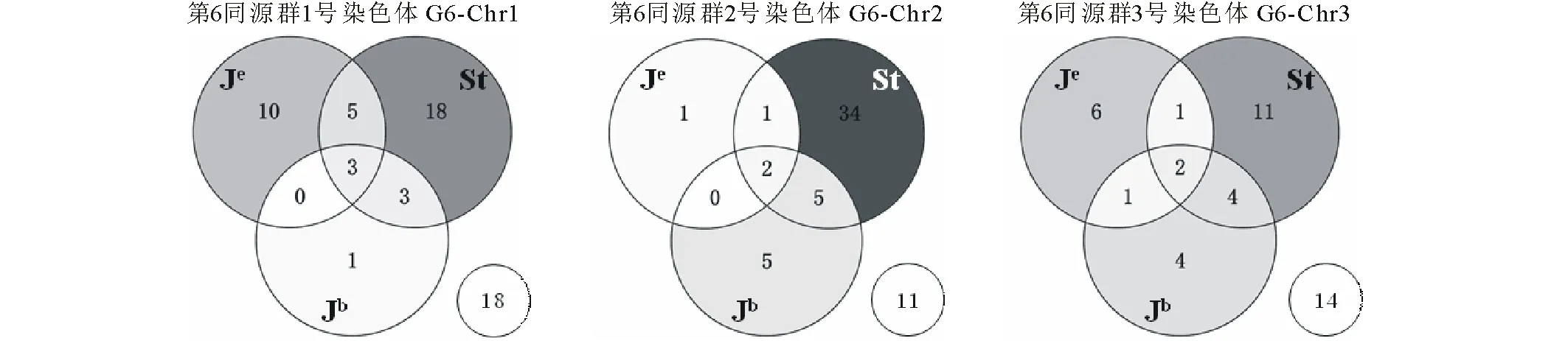
图3 第6同源群特异标记在Je/Jb/St基因组中的扩增Fig.3 Amplification of Thi-G6 specific markers in Je/Jb/St genomes图形颜色的深浅代表数目的多少。 The shade of the color of the Wayne diagram represents the number.
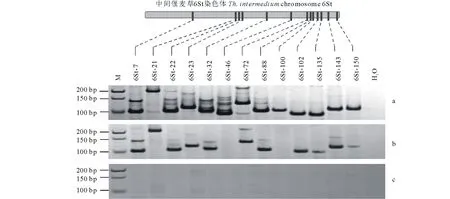
图4 “+/-”型Thi-6St特异标记在中间偃麦草Z1141(a)、小麦-偃麦草代换系F881(b)和中国春(c)中的扩增Fig.4 Amplification of “+/-” type Thi-6St specific markers in Th. intermedium Z1141 (a), wheat-Thinopyrum substitution line F881 (b) and Chinese spring (c) M: 指示带Marker ladder;6St-7~6St-150:中间偃麦草6St染色体“+/-”型特异标记 “+/-” type specific markers on Thi-6St.
3 讨论
自前苏联科学家Tsitsin[25]通过远源杂交首次获得小麦-中间偃麦草八倍体以来,利用附加系、代换系、易位系等材料将中间偃麦草的抗逆、优质等诸多农艺性状导入小麦,极大促进了小麦品种的遗传改良。在对远源杂交后代外源染色体的追踪过程中,原位杂交技术是最为直观和最有说服力的鉴定手段,但是其鉴定结果还不能精确到染色体具体位置,此外对于高代材料中的外源小片段、特别是对于隐性渗入系[26-27]尚无法鉴别,因此需要分子标记的辅助鉴定。
本研究基于组装出的中间偃麦草基因组序列开发了一套STS标记,并从第6同源群上的253个标记中筛选出107个多态性标记,即因中间偃麦草和普通小麦基因组序列的部分同源性而扩增出不同长度产物的特异标记,该比例(42%)与利用小麦第6同源群SSR标记来验证小麦-中间偃麦草间多态性的实验结果(41%)一致[28];另外本研究还筛选出53个只在中间偃麦草中有扩增产物的“+/-”型特异标记,丰富了鉴定标记的数量。分析这些特异标记所在染色体的基因组组成发现,G6-Chr2特异标记在拟鹅观草中的特异性最高,因此最容易鉴定出其为6St;G6-Chr1上的标记在Je+Jb中特异的共有11个,在拟鹅观草中特异的有18个,占62%,而 G6-Chr3上的标记在拟鹅观草中特异的占52%,两者无明显差异,因此尚不能确定 G6-Chr1和 G6-Chr3的染色体组成,今后需使用中间偃麦草6Js或6J 在小麦中的附加系、代换系材料加以鉴定。6Js与6J 不易区分可能是由于中间偃麦草 Js染色体组与 J 染色体组非常相近,只在其染色体着丝粒区域包含部分 St 染色质[7]所致,此外,Thi-G6特异标记中还有43个(27%)未能在任何一个祖先种中扩增出特异条带,意味着中间偃麦草在由祖先种自然杂交形成多倍体后,其基因组经历了较深层次的重组,逐渐向新物种的方向演化,这个现象在自然界普遍存在[29]。
由于 J 组和 St 组是两个麦族基础基因组[13,30],因此,中间偃麦草特异标记还可以用于鉴定含有 J 或 St染色体的其他麦族物种。利用53个“+/-”型Thi-G6特异标记对一份小麦和长穗偃麦草(2n=10x=70,JJJJJJJsJsJxJx)的代换系材料 X005(6Js/6B)[31]进行验证,结果显示 G6-Chr1-46、70、107和181等9个标记可以鉴定出 X005中的外源染色体6Js。本研究下一步将对其余6个同源群的分子标记进行筛选,开发整套中间偃麦草特异标记,这将为中间偃麦草甚至其他麦族近缘物种的鉴定提供较为快速和经济的分子检测手段,这些标记既可用于麦族物种的分类和进化研究,也可用于小麦育种工作中的遗传背景改良。
4 结论
本实验基于组装的中间偃麦草序列共开发2019对STS标记,其中第6同源群有253对,经过筛选获得160对特异标记,包含58个G6-Chr1标记、59个G6-Chr2标记和43个G6-Chr3标记。其中G6-Chr2为6St染色体,利用6St上的13个“+/-”型特异标记,可验证小麦-偃麦草代换系材料F881中导入的外源6St染色体。
