黄土台塬不同林型土壤主要温室气体通量特征
2019-04-22刘梦云张萌萌杨静涵曹润珊
张 杰 ,刘梦云 *,张萌萌 ,杨静涵 ,曹润珊
(1西北农林科技大学资源环境学院,陕西杨凌712100;2农业部西北植物营养与农业环境重点实验室,陕西杨凌712100;3农业部农业环境重点实验室,北京100081)
气候变暖是当今人类生存面临的最大环境问题,其首要原因归因于温室气体排放。二氧化碳(CO2)、甲烷(CH4)和氧化亚氮(N2O)作为3种主要温室气体,其排放量对温室效应的贡献十分明显[1]。有研究[2-3]指出,大气中每年有5%~20%的CO2、15%~30%的CH4、80%~90%的N2O来源于土壤,土壤中的微生物将有机质分解为无机态的碳和氮,在好氧条件下无机碳多以CO2形式进入大气,而在严格厌氧条件下则以CH4形式排向大气。全球一半以上的N2O来自于土壤的硝化与反硝化过程[4-6]。森林作为地球上分布最广的陆地生态系统[7],其植被可以通过改变土壤温湿度、根系呼吸、光合产物及小气候等途径影响土壤呼吸及各种温室气体的排放,但该效应在不同植被尤其是不同林型组成之间差异很大[8],因此,探究区域内不同林型组成之间土壤温室气体排放量的动态变化特征和及其对各环境因子的响应,对于合理预测植被的C、N排放源或吸收汇功能以及对大气碳氮循环的影响具有重要的科学意义。
黄土台塬毗邻西北黄土高原南部边缘地带,生态环境脆弱,水土流失严重,为防止土壤侵蚀,历年来塬坡防护林一直以水土保持、发挥生态效益为主要目标,从二十世纪六七十年代开始当地大面积种植刺槐纯林,随后引入松类,但由于纯林遗传性单一,林分结构呈不稳定状态,对外界环境适应性差,因此具有很大的弊端,为了解决这一问题,当地相继引进了针叶或阔叶等乔木以及沙棘等灌木林,开展了营造混交林试验研究,因此,黄土台塬区现存树种多为人工纯林或混交林。为了探明该区域不同林型组成下土壤CO2、CH4和N2O的时间变化特征以及对各类因素的响应,本研究以黄土台塬4种典型林地及其混交林共7种林型为研究对象,采用静态箱-气相色谱法对CO2、CH4和N2O通量进行测定,分析不同林型组成下土壤温室气体排放差异,阐明土壤温室气体与土壤温度和土壤含水量之间的关系,以期为评价和预测我国黄土台塬地区不同林型组成碳、氮的源/汇功能提供实测数据。
1 材料与方法
1.1 研究区概况
试验于陕西省咸阳市永寿县马莲滩林场(108°05′22.6″E~108°05′39.3″E;34°47′58.4″N~34°48′42.7″N)进行,该林场属于国家科技攻关项目“黄河中游黄土高原区(永寿)水土保持型植被建设技术研究与示范”实验基地,试验区占地面积为26.27 km2,其中,山地、沟壑面积 8.20 km2,塬面 10.87 km2,该区主要土壤类型为黑垆土,属温带大陆性季风气候,干旱指数为1.73~1.80,平均气温10.8℃,极端最高温38.8℃,平均降水量601.6 mm。由于研究区以台塬、沟壑区为主,生态环境极端脆弱,为解决当地恶劣的生态问题,该林场实行了封山育林政策,进行不同植被类型、不同混交方式与栽培模式的试验研究,并采用自然恢复的管理措施,后期人为干预较少。目前,林场森林植被以天然次生林(侧柏等)、人工林(刺槐、油松等)以及灌木(沙棘等)为主。
1.2 试验设计
该试验于2015年4月至2016年3月进行,选取晴朗天气作为采集日,时间间隔为一个月左右,选取当天11:00—15:00采集气体,以降低早晚温差对土壤呼吸速率的影响[9],将研究区7种林型分成3个实验小组,a组为灌木林(沙棘)、乔木林(刺槐)及乔灌混交林(沙棘刺槐混交林),b组为阔叶林(刺槐)、针叶林(侧柏)及针阔混交林(刺槐侧柏混交林),c组为灌木林(沙棘)、阔叶林(刺槐)、针叶林(油松)及乔灌-针阔组合型混交林(沙棘刺槐油松混交林),各林型林龄约为16~20 a,林下植被以关中地区自然恢复典型植被(蒿类、赖草、羊茅、蔷薇、悬钩子等)为主,每种林型选择3个样本作为对照,取其平均值作为当月该林型土壤的土壤呼吸速率。采样点情况如表1所示。
1.3 土壤基本理化性质测定
土壤全氮(TN)采用开氏法消解,K2300型全自动定氮仪测定;土壤有机碳(SOC)采用重铬酸钾-硫酸消化法测定;溶解性有机碳(DOC)采用0.5 mol·L-1K2SO4溶液浸提过筛风干土,经振荡机振荡后过滤,滤液用TOC自动分析仪测定有机碳,即为DOC;碱解氮采用碱解扩散法测定;NH+4-N、NO-3-N采用KCl溶液浸提新鲜土样,经振荡机振荡后过滤,滤液用流动分析仪测定;土壤pH采用pH计测定;土壤容重采用环刀法测定。
1.4 土壤温室气体排放通量测定
本实验采用静态箱(箱体材质为PVC,直径15 cm,桶高25 cm)气相色谱法收集温室气体,在安置箱体前将基座内的地表植物齐地剪掉,最大程度减少对地表凋落物的扰动,降低对土壤表层的破坏。将静态箱置于地下10 cm左右,防止因土壤松动造成气体泄漏,依序在第0、10、30、60、90、120 min采集气体,将10 mL医用针管插入带有橡皮塞箱盖的静态箱中采集气体,在此期间,用地温计测定采样点地下5 cm和20 cm处的土壤温度;同时,采集0~5、5~20 cm土壤样品用于土壤湿度的测定,用气相色谱仪7890A测定气体中CO2、CH4和N2O的体积分数,温室气体排放速率计算公式为[10]:

式中:F为温室气体通量,mg·m-2·h-1;dc/dt为气体浓度随时间变化的斜率(c为t时刻被测气体浓度,t是时间);M为被测气体的摩尔质量,g·mol-1;V0为标准状态下的气体摩尔体积,22.41 L·mol-1;P为采样点的大气压;P0为标准大气压,101.325 kPa;T为箱内温度,K;T0为绝对温度,273.15 K;H为采样箱的高度,cm。
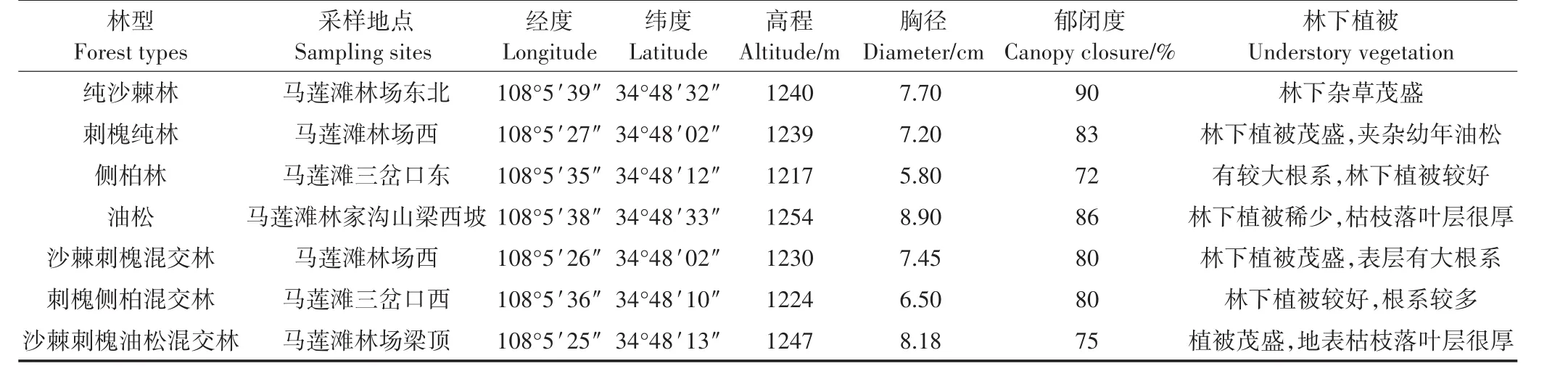
表1 7种林型组成基本情况Table 1 Basic status of sampling sites under different forest types
1.5 温度敏感性指数(Q10)
用Q10值表示温度增加10℃时土壤呼吸速率增加的倍数,并采用指数关系模型进行计算[38]:
Rs=a·ebT1,Q10=e10b式中:Rs为土壤呼吸速率,mg·m-2·h-1,T1为土壤温度,℃;a,b为待定参数。
1.6 数据分析与处理
所有数据均在Excel 2010和SPSS 21.0统计分析软件中进行,利用Pearson相关分析和单因素方差分析计算不同林型和季节土壤温室气体的差异,绘图均在Origin8.1软件中完成。
2 结果与分析
2.1 不同林型土壤温度和土壤含水量的季节性动态特征
森林土壤温湿度季节变化明显(图1),各林型土壤温度均表现为夏高冬低的规律,不同林型组成之间土壤温度差异很小,对比发现,冬季混交林的土壤温度普遍高于纯林,这是因为冬季混交林枯枝落叶多,对土壤起到一定的保温作用,且各林型上层土壤温度普遍低于下层土壤温度,这种现象与当地冰雪覆盖周期有关,而夏季则正好相反,这与夏季混交林郁闭度高、抵挡了部分太阳辐射到达地面有关。不同于土壤温度的变化规律,各林型土壤含水量在生长季(5—10月)变化平缓,而非生长季(11—来年4月)土壤含水量波动明显。7种林型之间土壤含水量较土壤温度差异明显,其中,沙棘年均土壤含水量在各林型中居于首位,而其他几种纯林则普遍低于混交林。
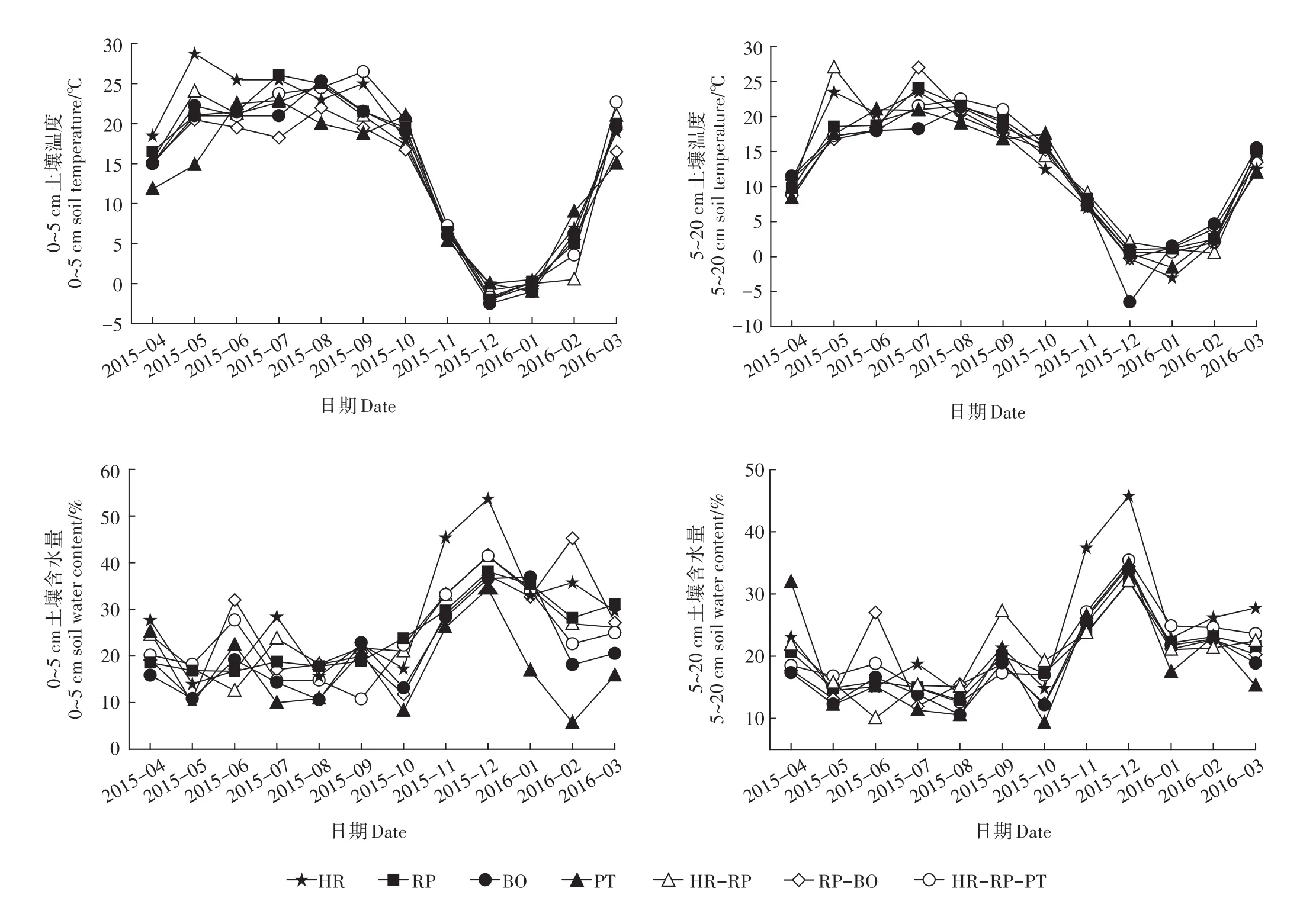
图1 7种林型0~5、5~20 cm土壤温度和土壤含水量的变化Figure l Seasonal changes in soil temperature and soil water content at 0~5 cm and 5~20 cm depths for the seven forests
2.2 不同林型土壤CO2通量的季节变化特征
研究期间,各林型土壤CO2排放通量夏季高、冬季低(图2),与土壤温度的变化趋势相似,且每年3月份随冻结土壤解冻,各林型土壤CO2通量会出现短暂回升;同时,混交林土壤CO2排放峰值在时间上往往晚于纯林1~2个月,这可能与各林型对土壤温湿度的适应范围有关。不同林型年均CO2排放通量表现为:沙棘刺槐混交林(130.73 mg·m-2·h-1)>刺槐(108.31 mg·m-2·h-1)>沙棘(107.78 mg·m-2·h-1)>刺槐侧柏混交林(81.95 mg·m-2·h-1)>侧柏(63.71 mg·m-2·h-1)>沙棘刺槐油松混交林(61.52 mg·m-2·h-1)>油松(51.33 mg·m-2·h-1)。此外,土壤 CO2排放通量与林型有关(表3),其中,a组沙棘刺槐混交林的土壤CO2通量在生长季显著(P<0.05)高于沙棘(34.01%)和刺槐(37.04%),而在非生长季则显著(P<0.05)低于沙棘(19.52%)和刺槐(26.29%);b组刺槐侧柏混交林的年均土壤CO2排放通量居于两种纯林之间,尤其在生长季显著(P<0.05)低于刺槐(13.56%)且高于侧柏(40.97%);c组沙棘刺槐油松混交林在生长季/非生长季均显著(P<0.05)低于沙棘(41.36%/47.95%)和刺槐(40.03%/52.33%),与油松差异不明显(P>0.05)。由此看来,不同林型组成之间土壤CO2排放通量存在一定程度的差异,在本研究中,沙棘刺槐混交林的土壤CO2通量大小在生长季和非生长季表现出相反的规律,刺槐侧柏混交林由于种间矛盾的存在,中和了其纯林的土壤CO2排放特征,而作为乔灌混交和针阔混交组合型的复杂林型,沙棘刺槐油松混交林土壤CO2排放量明显降低。
2.3 不同林型土壤CH4通量的季节变化特征
7种林型土壤CH4通量均为负值(图3,表3),表明研究区各林型土壤对CH4主要起吸收作用。从季节变化来看,各林型土壤CH4吸收生长季略高于非生长季,同时,在非生长季期间各林型的土壤CH4通量波动明显。研究中各林型年均CH4吸收通量表现为:油松(60.82 –g·m-2·h-1)>沙棘(56.74 –g·m-2·h-1)>刺槐(53.23–g·m-2·h-1)>沙棘刺槐混交林(47.44 –g·m-2·h-1)>刺槐侧柏混交林(47.12 –g·m-2·h-1)>沙棘刺槐油松混交林(38.23–g·m-2·h-1)>侧柏(36.67 –g·m-2·h-1)。各混交林土壤CH4吸收通量在生长季略低于相应纯林,但差异并不显著(P>0.05);对比非生长季各林型土壤CH4吸收通量可以看出,a组沙棘刺槐混交林土壤CH4吸收通量低于刺槐(3.50%),且显著(P<0.05)低于沙棘(28.70%);b组中刺槐侧柏混交林与刺槐的土壤CH4吸收通量差异不大(P>0.05),但显著(P<0.05)高于侧柏(79.65%),出现这种现象的原因可能与侧柏的浅根性、土壤易氧化有关;c组沙棘刺槐油松混交林土壤CH4吸收通量显著(P<0.05)低于沙棘(48.11%)、刺槐(37.00%)和油松(47.98%)。综合对比3组不同林型之间土壤CH4通量变化特征可以发现,混交林与纯林相比,其土壤对CH4的吸收作用较弱;但3种组合形式的混交林土壤CH4吸收能力依次为:沙棘刺槐混交林>刺槐侧柏混交林>沙棘刺槐油松混交林。
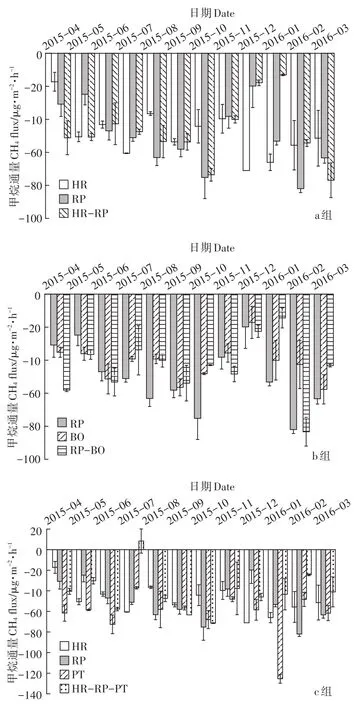
图3 不同林型土壤CH4通量的变化特征Figure 3 Seasonal changes in soil CH4fluxes for the seven forests
2.4 不同林型土壤N2O通量的季节变化特征
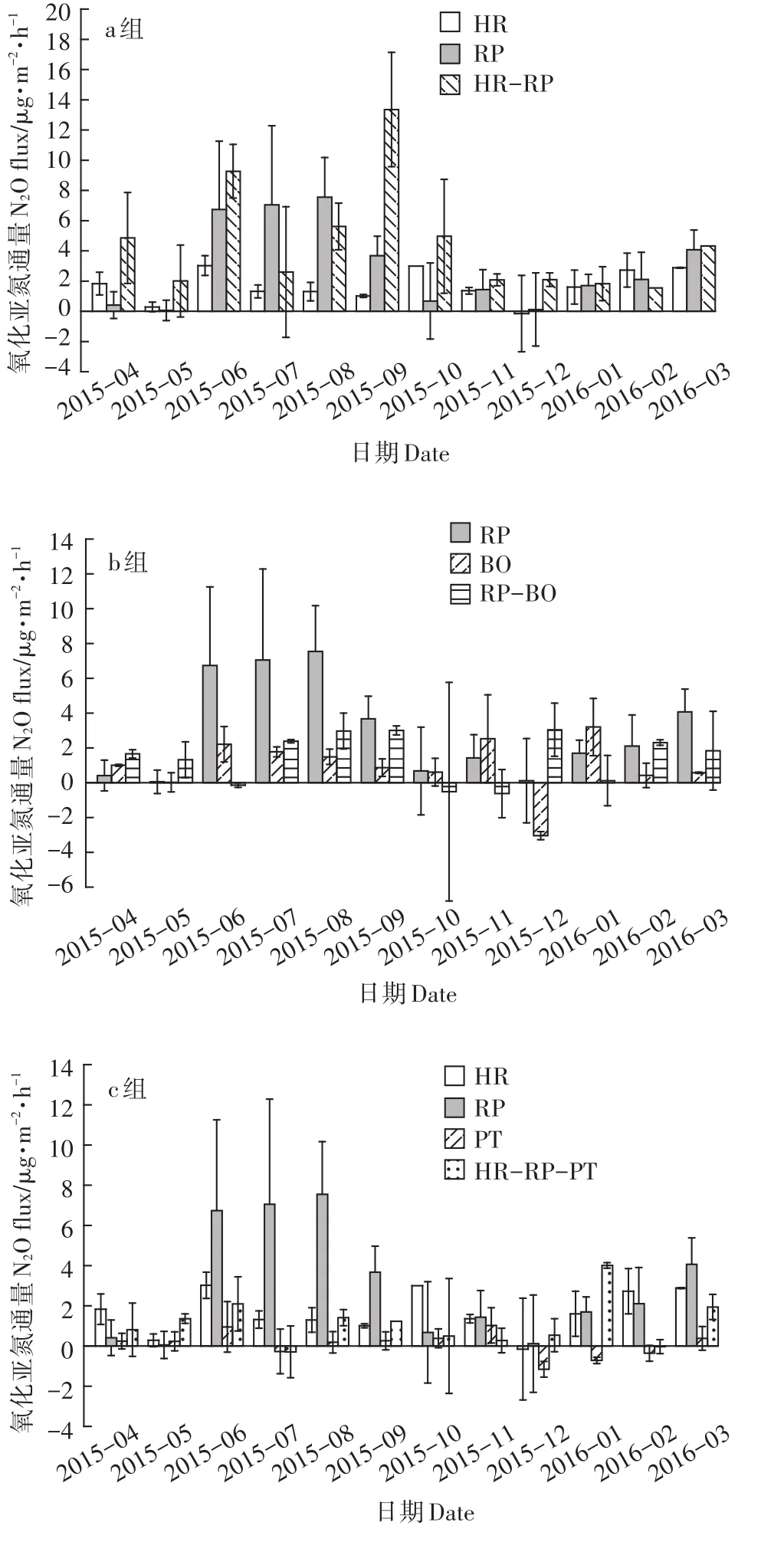
图4 不同林型土壤N2O通量的变化特征Figure 4 Seasonal changes in soil N2O fluxes for the seven forests
各林型土壤N2O通量水平较低(图4),夏高冬低是几种林型土壤N2O排放通量变化的普遍规律,在个别月份出现N2O的净吸收现象。各林型土壤年均N2O通量表现为:沙棘刺槐混交林(4.42 –g·m-2·h-1)>刺槐(4.00 –g·m-2·h-1)>刺槐侧柏混交林(1.55 –g·m-2·h-1)>沙棘(1.52 –g·m-2·h-1)>沙棘刺槐油松混交林(1.05–g·m-2·h-1)>侧柏(0.72 –g·m-2·h-1)>油松(-0.06 –g·m-2·h-1)。不同林型之间N2O通量差异显著(表3),从a组来看,沙棘刺槐混交林在生长季显著(P<0.05)高于沙棘(407.26%),略高于刺槐(10.35%);而在非生长季,3种林型N2O通量差异并不显著(P>0.05)。其次,通过b组比对发现,刺槐侧柏混交林土壤N2O通量居于刺槐和侧柏之间,生长季显著(P<0.05)低于刺槐(73.68%),非生长季则显著(P<0.05)高于侧柏(492.59%);c组沙棘刺槐油松混交林的土壤N2O通量与沙棘之间差异不显著(P>0.05),但显著(P<0.05)低于刺槐(生长季/非生长季:81.15%/54.15%),与油松的差异仅表现在非生长季,油松的土壤N2O排放通量较低但季节变化明显,尤其在冬季油松的土壤N2O通量出现负值,此时其土壤明显作为N2O的库,因此显著(P<0.05)低于其混交林(138.1%),其他几种林型仅在极低气温的月份出现负值但并不持久。总体来看,各林型土壤N2O通量表现出与土壤CO2相似的规律,即沙棘刺槐混交林土壤N2O排放量在3种组合形式的混交林中居于首位,刺槐侧柏混交林居中,而沙棘刺槐油松混交林土壤N2O排放量最低。

表3 不同林型土壤CO2、CH4和N2O通量比较(平均值±标准差)Table 3 Comparison for the soil fluxes of CO2,CH4and N2O in seven forests(Mean±SD)
2.5 温室气体通量与土壤温度和土壤含水量的关系
各林型土壤CO2排放与土壤温度均呈显著(P<0.05)或极显著(P<0.01)指数正相关(图5),但不同林型及不同土层深度的相关性存在差异。其中,沙棘刺槐混交林和刺槐侧柏混交林土壤CO2与土壤温度的相关性与其纯林相比较低,而沙棘刺槐油松混交林土壤CO2与温度的相关性则高于其纯林;同时,5~20 cm深度的土壤CO2与土壤温度的相关性整体高于0~5 cm土层。各林型土壤表面CH4通量与土壤温度呈线性负相关,但相关性较低,仅侧柏的土壤CH4通量与土壤温度具有极显著(P<0.01)相关性,且在0~5 cm深度上的相关性高于5~20 cm土层;沙棘刺槐混交林仅在0~5 cm土层深度上与土壤温度呈显著(P<0.05)相关,其他林型无明显相关性。在土壤N2O排放中,仅刺槐和油松与土壤温度呈线性显著(P<0.05)正相关,且刺槐土壤N2O在5~20 cm深度上与土壤温度的相关性高于0~5 cm,而油松土壤N2O仅在5~20 cm深度上与土壤温度相关,在0~5 cm深度上未表现出相关性。
土壤含水量与各林型土壤CO2表现为负相关性(图6)。其中,沙棘刺槐油松混交林土壤CO2与土壤含水量相关性最高,且在5~20 cm深度上相关性高于0~5 cm土层;刺槐和油松的土壤CO2与土壤含水量的相关性在0~5 cm深度上高于5~20 cm土层;而沙棘刺槐混交林土壤CO2仅在0~5 cm深度上与土壤含水量显著(P<0.05)相关,其他林型土壤CO2与土壤含水量相关性不显著(P>0.05)。各林型土壤CH4通量与土壤含水量的关系相对复杂,其拟合曲线是开口向上的抛物线,即随土壤含水量的增加,土壤对CH4的吸收量先增加后减少,且这种现象只在侧柏和沙棘刺槐混交林0~5 cm土层深度中表现显著(P<0.05)。而各林型土壤N2O对土壤含水量的响应较弱(P>0.05)。
3 讨论
3.1 不同林型土壤表面CO2通量排放特征
在土壤温湿度的影响下,土壤呼吸速率具有明显的季节变化[11-13]。本研究中,各林型土壤CO2通量均为夏高冬低,这是因为夏季土壤温度高,水分条件适宜,植被生长旺盛,而冬季与之相反,且黄土台塬冬季降雪频繁,温度比同纬度其他区域更低,冰雪覆盖下表层土壤易形成季节性冻土,阻碍土壤中CO2向大气扩散。研究表明,针叶林、乔灌-针阔组合型混交林土壤CO2排放量较低,其次为针阔混交林,而乔灌混交林及其纯林土壤CO2通量较高。这与Raich等[14]的研究结果相符。各林型土壤CO2排放特征与不同森林类型之间凋落物数量、有机质含量、氮素数量、pH及土壤容重等差异有关[15],在本研究中,针叶林(油松)土壤全氮、有机碳含量均低于其他林型(表2),相关分析显示(表4),土壤CO2通量与土壤有机碳呈显著(RSOC/5~20=0.85,P<0.05)正相关,与全氮呈极显著(RTN/0~5=0.90,P<0.01)正相关,这与耿远波等[16]的研究一致,土壤有机碳为异氧呼吸提供物质基础,而土壤呼吸与反硝化作用有较好的相关性,这与氮能刺激植物初级生产为土壤呼吸提供更多底物有关[13],同时,氮含量的高低又会影响C/N,但目前对于C/N与土壤CO2的关系还不明确,本研究中,两者有一定的正相关关系,但未达显著性水平(P>0.05)。在本文中,土壤pH对土壤CO2的影响并不显著(P>0.05),有研究[17]指出pH并不是造成土壤温室气体排放差异的原因,而是通过酸化累计过程影响土壤碳、氮等养分从而间接影响CO2排放。此外,通过枯枝落叶的分解向大气释放CO2是土壤碳循环的一个重要环节[15],与其他林型相比,针叶分解速度较慢。有研究[18]指出刺槐和油松之间存在异株克生现象,刺槐作为速生大冠阔叶树种,当用其作油松的伴生树种时,超过一定的比例会压制油松的生长,因而沙棘刺槐油松混交林具有明显的低CO2排放特征。而乔灌混交林及其纯林土壤养分含量较高,又具备较低的土壤容重,故其土壤CO2排放速率较快。针阔混交林因其合理的林分结构,土壤肥力较好,土壤呼吸速率在各林型中处于中等水平。进一步分析混交林及其构成纯林之间土壤CO2排放差异可能与土壤温湿度有关,土壤温度和水分是森林土壤呼吸的重要环境因子,但两者的影响机制有所差异[12,19],以研究区刺槐侧柏混交林为例,植被在生长季温度条件适宜的情况下,土壤含水量极易成为土壤呼吸的限制因素,研究结果表明,刺槐侧柏混交林在生长季土壤含水量显著(P<0.05)低于刺槐且高于侧柏,这一现象与其土壤CO2排放特征具有相似性。受地表大气温度影响,0~5 cm土壤温度与土壤CO2的相关性低于5~20 cm土壤温度,通过计算各林型Q10值发现(表5),灌木林和针叶林土壤呼吸对0~5 cm土壤温度的敏感性低于5~20 cm土层深度,这与众学者[20-21]的研究一致。相关研究[22]表明,Q10值与底物供应的季节变化存在正相关性,阔叶林凋落物累积量常常较纯林多,对照0~5 cm土层各林型发现,落叶阔叶林和混交林Q10值高于灌木和针叶林。大量研究[19,23]表明土壤含水量与土壤CO2呈正相关,在本研究中,各林型土壤CO2与土壤含水量为负相关,这与前人研究结果不同,造成本研究结果与以往研究不同的原因可能是研究区气候条件复杂,尤其是水、热不同期,土壤含水量升高时土壤温度反而降低,低温造成土壤微生物活性降低,土壤呼吸作用减弱,同时较高的含水量造成土壤孔隙度变小,进而抑制了土壤CO2的排放。

图5 温室气体与0~5、5~20 cm土壤温度的关系Figure 5 Relationship between soil greenhouse gases and soil temperature at 0~5 cm and 5~20 cm depths for the seven forests

图6 温室气体与0~5、5~20 cm土壤水分的关系Figure 6 Relationship between soil greenhouse gases and soil water content at 0~5 cm and 5~20 cm depths for the seven forests
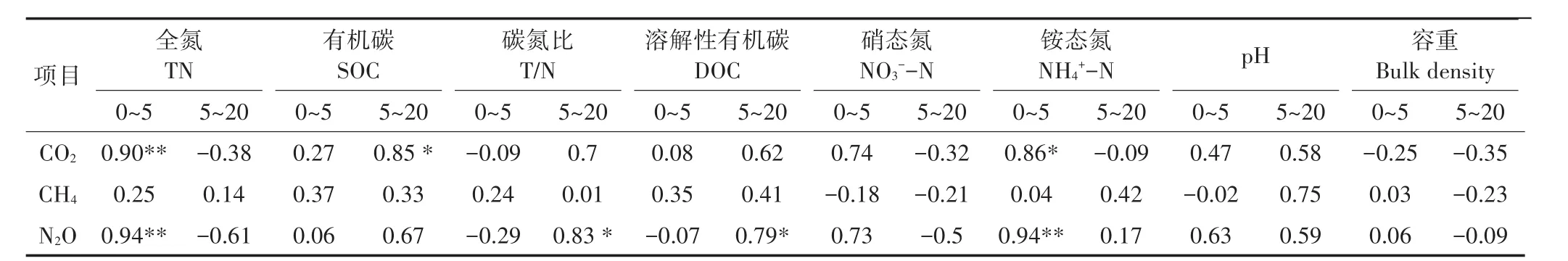
表4 森林温室气体排放速率与土壤理化性质的相关系数Table 4 Correlation coefficient between greenhouse gas fluxes rate and physicochemical properties at different soil depths

表5 不同林型土壤呼吸的温度敏感性指数(Q10)Table 5 Temperature-sensitive index(Q10)of soil respiration at different soil depths for the seven forests
3.2 不同林型土壤表面CH4通量排放特征
研究期间,各林型土壤均为CH4汇,这与众学者[24-25]研究结果相符。研究表明,各林型土壤CH4通量在生长季无显著(P>0.05)差异,但在非生长季差异显著(P<0.05),这是由于研究区在非生长季内土壤含水量较高,而CH4氧化主要受土壤通气状况的影响,低温高湿的土壤环境不利于土壤CH4氧化。同时,土壤冻融交替过程也会影响土壤通气状况和氧化还原条件[26],各林型土壤CH4吸收能力在2月份急剧下降,这是因为冬季冻结土壤营造的厌氧环境下产生了大量CH4气体,土壤解冻后会释放其储存的CH4。有研究[27]指出,CH4排放与易矿化碳存在明显线性关系,DOC作为微生物生长的速效基质,能显著提高CH4生成量[16,27],尽管本文中两者相关性并不显著(P>0.05),但相比土壤全氮、有机碳含量及C/N,该土壤养分含量对 CH4通量的解释率较高(RDOC/5~20=0.41)(表 4)。pH对土壤CH4排放通量的影响主要通过改变土壤微生物和酶实现,研究发现森林土壤甲烷氧化菌最适土壤pH约为5.0~6.5[28],研究区各林型土壤pH值偏弱碱性,对土壤CH4氧化的影响较低。无机氮对森林土壤CH4吸收的贡献目前还不明确,很多学者在研究中均指出NH+4浓度对CH4吸收具有抑制作用,并将其解释为NH3与CH4在甲烷单氧化酶上的竞争[29],但在本研究中土壤CH4排放与NH+4相关性不显著(P>0.05),这可能与研究区土壤NH+4-N含量较低,植被生长成为氮素竞争的优先者有关。以非生长季乔灌-针阔组合型混交林及其纯林土壤CH4吸收为例,对比纯林与混交林土壤温度发现,各林型之间非生长季土壤温度差异并不明显(P>0.05),而混交林土壤含水量则在其构成纯林之间浮动,由此可见,土壤湿度对不同林型土壤CH4吸收具有潜在影响,但其影响机制较复杂,本研究发现,土壤含水量与森林土壤CH4通量并不是简单的线性关系,在植被生长过程中对土壤水分的生理需求较大,土壤水分的微弱变化就能引起土壤微生物活性的剧烈变化,同时,不同林型在生长过程中所需土壤湿度的最适阈值不同,即当土壤含水量处于植物生长最适区间时,有助于土壤对CH4的吸收,而当土壤含水量过高或过低时,均会抑制CH4氧化速率。这与以往研究[15]结果一致。相关分析显示,土壤温度对针叶林(侧柏)和乔灌混交林土壤CH4吸收具有促进作用,而其他几种林型土壤CH4通量与土壤温度相关性不显著(P>0.05),这是因为温度的影响只有在特定水分条件下才能表现出来,而各林型之间土壤含水量存在一定差异;同时,0~5 cm深度的土壤温度与针叶林土壤CH4相关性较高,这可能与0~5 cm深度的土层中能被微生物分解的有机物质含量高有关。Hansen等[2]在其研究中也证实了CH4氧化作用主要发生在表层土壤之中,随土层深度增加土壤CH4的氧化作用逐渐减弱。
3.3 不同林型土壤表面N2O通量排放特征
各林型土壤N2O排放通量与已有研究[30]相比排放水平较低,这是由于土壤N2O主要来源于外界人为施氮,而研究对象位于封山育林区,人为影响较小,故土壤N2O排放量较低。崔骁勇等[19]在其研究中指出,受高温季节的影响,土壤N2O排放会达到高峰,这与本研究中各林型土壤N2O排放整体呈夏季高、冬春低的规律较为一致。不同林型组成之间由于土壤养分、土壤质地及土壤温湿度等理化指标不同,进而影响到土壤硝化和反硝化过程,造成各林型土壤N2O的排放差异[31]。近年来众学者将氮素水平作为研究土壤N2O的重点,徐慧等[30]在其研究中指出土壤N含量是影响土壤N2O的重要因素,同时,土壤中DOC及NO-3-N作为土壤硝化和反硝化过程的基质,将明显影响N2O的排放[32],充足的DOC会刺激土壤中微生物的活性,加快土壤中O2消耗,为反硝化过程创造低氧环境;NO-3-N作为反硝化过程的底物,对反硝化强度有直接影响,但存在影响界限,即当NO-3-N浓度大于25 mg·kg-1时,土壤反硝化速率不受NO-3-N含量的影响,上述研究结果在本研究中也得到了证实(表4),即土壤全氮和DOC与土壤N2O排放呈极显著或显著(RTN/0~5=0.94,P<0.01;RDOC/5~20=0.79,P<0.05)相关,而NO-3-N与土壤N2O相关性并不显著(P>0.05);同时,本研究C/N值普遍低于25~30∶1(土壤微生物适宜有机质C/N[33]),微生物活性较强,促进土壤N2O的产生。李海防等[34]在其研究中曾指出固氮植物能直接通过根瘤菌参与下的硝化、反硝化作用或间接增加N量促进N2O的排放。在本文中,沙棘属于非豆科固氮树种,刺槐属于豆科固氮树种[35],根部具有固氮作用的根瘤菌,极大提高了混交林的土壤N含量,针阔混交林作为固氮和非固氮树种的混交林,提高了非固氮树种土壤肥力。而针叶林作为非固氮树种,其土壤N含量较低,在非生长季,针叶林(油松)的土壤N2O排放通量为负值,说明在一定条件下土壤对N2O具有吸收作用,此结论与前人[7,36]研究结果一致;此外,在冬季冰雪覆盖条件下,厌氧环境的形成也可能会使土壤中的N2O继续转化为N2,进而出现排放负值。土壤pH对硝化和反硝化过程的影响较复杂,在本研究中各林型土壤pH值(7.84~8.24)接近,与土壤N2O之间没有明显的相关性,对研究区各林型土壤N2O排放差异并不构成影响。除刺槐和油松外,各林型土壤N2O通量尽管表现出随温度升高和湿度下降而增大的趋势,但并不显著(P>0.05),考虑到研究区N2O排放通量水平较低,水热交替变化,无益于硝化和反硝化作用产生N2O,而5~20 cm深处的土壤温度对刺槐和油松土壤N2O的影响较高,这可能与两种植被本身的深根性、喜干冷气候有关。
4 结论
(1)黄土台塬7种林型土壤均为CO2和N2O的排放源,CH4的吸收汇。
(2)不同林型土壤温室气体季节排放或吸收存在差异,在生长季,乔灌混交林土壤CO2通量显著高于纯林,针阔混交林土壤CO2通量居于针、阔叶纯林之间,而乔灌-针阔组合型混交林的土壤CO2通量则低于阔叶林高于针叶林,在非生长季,3种混交林均低于其纯林;在土壤CH4吸收中,3种混交林的土壤CH4吸收能力均低于其纯林;在土壤N2O排放中,乔灌混交林土壤N2O排放能力高于其纯林,针阔混交林居于针、阔叶纯林之间,而乔灌-针阔组合型混交林低于阔叶林高于针叶林。3种组合方式的混交林土壤CO2、N2O排放和CH4吸收能力从高到低依次为:乔灌混交林>针阔混交林>乔灌-针阔组合型混交林。
(3)不同组合方式的混交林土壤温室气体与土壤温湿度的相关性不同,与纯林、乔灌混交林以及针阔混交林相比,乔灌-针阔组合型混交林土壤CO2与土壤温湿度的相关性最高,其土壤CO2排放量随土壤温湿度的升高而增加;森林土壤对CH4的吸收随土壤温度升高而增加,但对土壤含水量有一定的适应范围,当超过一定的阈值时,土壤对CH4的吸收能力会随之减弱;各林型土壤N2O与土壤温湿度相关性较低,仅针、阔叶纯林土壤N2O表现出与土壤温度的正相关性,其他林型与土壤温湿度均无显著相关性。
