驱动蛋白的能量转换过程
——从ATP分子结合到颈链对接
2019-04-20马翼龙吕丽娜耿轶钊
马翼龙,李 铁,吕丽娜,耿轶钊*,纪 青*
(河北工业大学a.生物物理研究所;b.理学院;c.省部共建电工装备可靠性与智能化国家重点实验室;d.电气工程学院,中国天津300401)
驱动蛋白是一种普遍存在于真核生物细胞中的马达蛋白,能够与微管蛋白特异性结合并沿着微管定向连续行走,将尾部所携带的细胞器或大分子货物输送到细胞中的指定位置[1~3]。驱动蛋白又是一种核苷酸酶,能够催化ATP分子水解,将ATP分子携带的化学能转化为沿微管定向行走的机械能[4]。驱动蛋白种类非常多,可以分为14个亚家族[5~6],本文主要讨论最常见的以二聚体形式沿微管正方向连续行走的驱动蛋白[7~8]。现在已知驱动蛋白主要的发力做功过程涉及的构象变化是连接其马达结构域(motor domain,头部)的由十几个氨基酸组成的颈链向驱动蛋白马达结构域对接的过程[9]。此过程发生在二聚体中处于行走方向前方的驱动蛋白头部,可以将后面的头部带到其下一个微管结合位点,以这种方式,驱动蛋白行走一步[7,10~15]。但是,颈链的对接过程无法自发发生,需要ATP分子与此驱动蛋白马达结构域特异性结合,即需要消耗一个ATP分子。所以,ATP分子与驱动蛋白马达结构域的结合是驱动蛋白能量转化和力产生过程的起始步骤[9]。ATP分子与驱动蛋白ATP结合位点的结合所产生的构象和能量变化最终引起了颈链对接。但是,通过驱动蛋白马达结构域的结构(图1)可以发现,驱动蛋白的核苷酸结合位点和颈链分别位于马达结构域的两侧,ATP分子的结合无法直接对颈链产生影响。所以,两者之间必然存在着能够传递力和构象变化的结构[8,16~17]。在驱动蛋白结构-功能关系的研究中,一个关键的问题是ATP分子的结合效果如何传递到驱动蛋白的颈链部分并激发其向头部的对接运动,也就是驱动蛋白如何将化学能转化为机械能。本文结合本课题组的研究结果,分三个部分对此问题的研究进展予以综述。
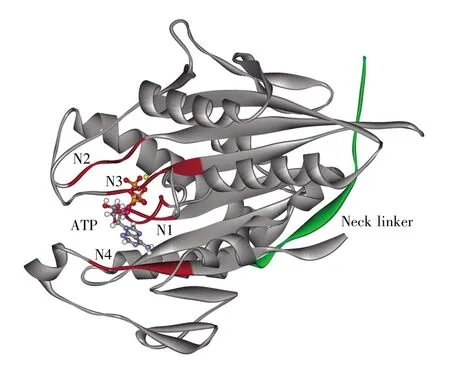
图1 驱动蛋白马达结构域的三维结构图中构成ATP结合位点的N1、N2、N3和N4模体标为红色。与马达结构域处于对接状态的颈链用绿色表示。ATP分子和Mg2+用球棍模型表示。Fig.1 The structure of kinesin motor domainThe ATP binding site motifs N1,N2,N3 and N4 are colored in red.The neck linker connected to motor domain is colored in green.The ATP molecule and Mg2+ion are shown in balland-stick models.
1 ATP分子与驱动蛋白ATP结合位点的相互作用
核苷酸酶的核苷酸结合位点都是高度保守的。Sablin等[18~19]对驱动蛋白、肌球蛋白和G蛋白的核苷酸结合位点进行了系统研究,认为它们都是由同一个祖先蛋白质进化得到的。与肌球蛋白和G蛋白相同,驱动蛋白的ATP结合位点也包含4个模体(motif):N1,也叫做P-loop;N2,也叫做switch-Ⅰ;N3,也是switch-Ⅱ的一个组成部分;N4模体(图1)[20~21]。在驱动蛋白中,N1模体主要与ATP分子的α、β磷酸基团以及和ATP分子螯合的Mg2+有相互作用;N2和N3模体主要与ATP分子的γ磷酸基团以及Mg2+有相互作用;而N4模体上的氨基酸残基与ATP分子的嘌呤环存在疏水堆积作用,并和N1模体一起形成与嘌呤环几何形状匹配的疏水区,对ATP分子起到稳定作用。
ATP分子携带4个单位负电荷,可以与驱动蛋白的ATP结合位点产生特异性结合,使得体系的总能量降低。目前,相关实验已经确认,ATP分子与驱动蛋白的ATP结合位点结合即可诱导颈链的对接[9]。此过程不需要ATP分子水解的能量。ATP分子水解释放的能量可能主要用来将驱动蛋白的结构恢复到与ATP分子结合之前的状态。ATP分子进入驱动蛋白ATP结合位点主要促使N2和N3模体产生较大的构象变化[22]。Parke等[23]根据Eg5蛋白(属于kinesin-5亚家族)不同化学状态的结构对比(3HQD[23]和1II6[24]),提出了ATP结合和水解的双水分子催化机制(two-water catalytic mechanism)。在此机制中,ATP分子进入ATP结合位点之后,N2模体的Arg234(3HQD晶体结构残基编号)和N3模体的Glu270之间形成盐键,从而将N2和N3模体连接在一起,使ATP结合位点闭合,隔绝了外界水分子对ATP分子和ATP结合位点残基之间相互作用的攻击,保证了ATP分子的水解环境[23](图2)。我们的分子动力学模拟研究发现,驱动蛋白ATP结合位点(N3)与微管之间有较强的相互作用,这对于稳定ATP分子的水解环境也有非常重要的作用[25]。
在关闭的ATP结合位点内,Mg2+和ATP分子固定了两个水分子。在ATP分子的水解过程中,一个水分子提供了水解所需的氢氧根离子,而另一个水分子则接受了第一个水分子剩余的氢离子并与Glu270产生相互作用,引起 Arg234和Glu270之间盐键的断裂。在此盐键被破坏之后,N2和N3模体之间的结合打开,为水解产物磷酸基团的释放提供了通道[23]。McGrath等[26]也用量子力学/分子力学杂交计算(QM/MM)的方法对Eg5催化的ATP分子水解过程进行了理论计算,得到了此过程中的能量转移路径。ATP分子水解完成之后,磷酸基团快速释放的过程会破坏驱动蛋白残基之间一系列的弱相互作用,从而使驱动蛋白马达结构域产生较大的构象变化。Nitta等[27~28]针对此问题开展了深入的研究工作,本文不再进行详细的讨论。

图2 ATP水解的双水分子催化结构图和相互作用网络图中结构取自3HQD[23]晶体结构。结晶时,核苷酸采用的是实验条件下无法水解的ATP类似物—ANP。ANP分子、两个水分子(W1和W2)以及相关氨基酸残基用球棍模型显示出来。此结构没有显示氢原子。Fig.2 The two-water catalytic mechanism of ATP hydrolysis and the interaction networkThe structure is obtained from crystal structure 3HQD[23].In the crystallization,the ATP molecule is replaced by its analogue-ANP,which cannot be hydrolyzed under experimental conditions.The ANP molecule,two water molecules(W1 and W2)and related residues are shown in ball-and-stick models.The hydrogen atoms are not shown in this structure.
Hwang等[29]通过长时间的分子动力学模拟计算发现,在ATP分子寻找其结合位点的过程中,马达结构域与ATP分子嘌呤环部分的疏水相互作用起主要作用。马达结构域上的N2模体、α0和loop5组成了一个漏斗型结构,可以捕获ATP分子并将其带到N1模体上。
虽然人们对ATP分子与驱动蛋白ATP结合位点残基之间的相互作用以及ATP分子的水解过程进行了大量的理论和实验研究,但是,到目前为止,还有两个关键的问题尚不十分清楚,即ATP分子如何在水环境中识别驱动蛋白的ATP结合位点;ATP分子通过什么路径以何种方式进入驱动蛋白的ATP结合位点。本课题组目前正在对这两个问题进行研究。
2 ATP分子结合引起的驱动蛋白马达结构域转动
根据前面的讨论可知,ATP分子结合到驱动蛋白ATP结合位点会引起相应残基的一系列构象变化,主要表现为ATP结合腔的关闭。但是,ATP结合位点的小的构象变化无法直接启动驱动蛋白颈链向马达结构域的对接,必须通过马达结构域其他部分对ATP结合位点的构象变化进行传递和放大才可以实现。
通过与肌球蛋白的类比,Vale等[8]提出驱动蛋白的α4螺旋(又叫switch-Ⅱ helix)可能与肌球蛋白的“relay helix”类似,在马达结构域的化学循环过程中像活塞连杆一样往复运动,在不同的核苷酸结合状态下处于不同位置。α4螺旋在ATP结合和水解过程中的位置变化可以传递到颈链,引起颈链向马达结构域的对接。但是,在不同核苷酸状态下高分辨率的冷冻电镜图像以及马达结构域和微管复合体的晶体结构都表明,α4螺旋是驱动蛋白的微管结合位点[10,12,30~32]。在ATP分子结合和水解过程中,α4螺旋和微管的相对位置没有变化,保持静止。所以,与肌球蛋白不同,α4螺旋在驱动蛋白结合和水解ATP分子过程中并没有大的位移。
通过对比空态和ATP结合态驱动蛋白马达结构域的结构,我们可以清楚地看到马达结构域在ATP分子结合的前后绕着α4存在一个转动(图 3)。Sindelar等[10,30,33~34]提出了从 ATP 结合位点到颈链的力学传递“跷跷板(seesaw)”模型(图4)。在此模型中,马达结构域Asn255(使用1MKJ[10]晶体结构残基编号)与微管的相互作用稳定了α4螺旋N端结构。而整个马达头部是一个以α4为底面,以中心β片为板面,以α4上的Leu258、Leu261和中心β片上的Phe82、Tyr84为支点的“跷跷板”结构。此结构在不同核苷酸结合态下的倾斜控制着马达结构域两侧ATP结合腔和颈链结合腔的打开与关闭。ATP分子与ATP结合腔的结合撬动了“跷跷板”的一端,使“跷跷板”向ATP结合位点一侧倾斜,从而打开马达结构域另一端的颈链结合腔,使颈链上的Ile325进入颈链结合腔,启动颈链对接。
最近几年,冷冻电镜技术分辨率的大幅提高以及马达结构域和微管复合体晶体结构的获得,使得人们可以更深入地从氨基酸水平讨论ATP分子结合的效果如何引起驱动蛋白颈链的对接。比如:Moores课题组[35]和 Sindelar课题组[36]各自用冷冻电镜方法对此问题进行了研究,而Lipowsky团队[37]和Kaplus团队[29]则采用分子动力学模拟的方法研究了ATP分子结合引起的驱动蛋白的构象变化。他们的研究都提到整个马达结构域可以分为3个亚结构域。ATP分子的结合可以引起亚结构域本身的构象变化以及亚结构域之间的相对位移。我们也通过粗粒化分子动力学模拟计算的方法模拟了从ATP分子结合到中心β片转动再到颈链对接的整个过程,找到了几个关键的中间状态并计算出了不同状态之间的自由能差[38]。Sindelar研究团队也对其“跷跷板”模型进行了改进,提出了“弯弓”(archery bow)模型[36]。在此模型中,马达结构域在不同核苷酸状态存在两个不同的缝隙。在空态,马达结构域ATP结合位点的N1模体和N2模体之间存在一个“核苷酸缝隙(nucleotide cleft)”,ATP分子可以通过此缝隙进入ATP结合位点。而ATP分子的结合会将N1模体向微管的方向拉动约4 Å的距离并与N3模体产生相互作用,进而使“核苷酸缝隙”关闭。此构象变化也引起α2螺旋和中心β片的转动,有利于N3模体和α4螺旋之间的“聚合物缝隙(polymer cleft)”打开。在此过程中,中心β片存在弯曲。N1模体和α2螺旋像弓箭的弓弦一样,而β3和中心β片组成了弓背。在空态时,N1模体与N3模体分离,“弓”处于放松状态。ATP结合之后,N1和N3模体之间存在相互作用,“弓”处于拉紧的状态。而α2与N1模体之间的拉力传递到β4以及中心β片上,引起中心β片的扭曲和转动,进而引起颈链向马达头部的对接。
此部分的研究是驱动蛋白能量转换机制研究最关键的内容,从实验到理论计算方面都有大量的研究结果。但是,到目前为止,还无法从原子水平上将整个构象变化的图景描述清楚,尚存在非常多的细节问题,需要更深入的研究。
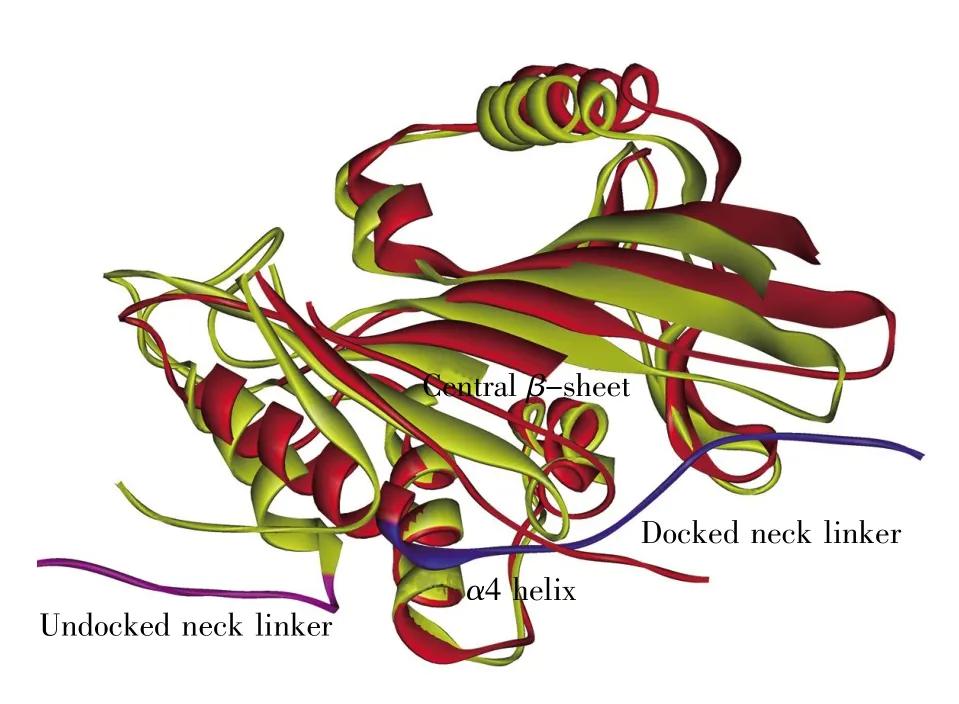
图3 驱动蛋白马达结构域在ATP分子结合前后的对比图空态(ATP分子结合之前)的马达结构域用黄色表示,ATP结合态的马达结构域用红色表示。处于非对接状态的颈链用紫色表示,处于对接状态的颈链用蓝色表示。为了清晰地显示中心β片绕α4螺旋的转动,图中隐去了驱动蛋白头部的 α1、α2 螺旋。Fig.3 Structure comparison of motor domain before and after ATP bindingThe motor domain structure before ATP binding is presented in yellow and that after ATP binding is colored in red.The undocked neck linker is colored in purple and the docked neck linker is colored in blue.To clearly show the rotation of central β-sheet around α4,the α1 and α2 helices are omitted in this figure.
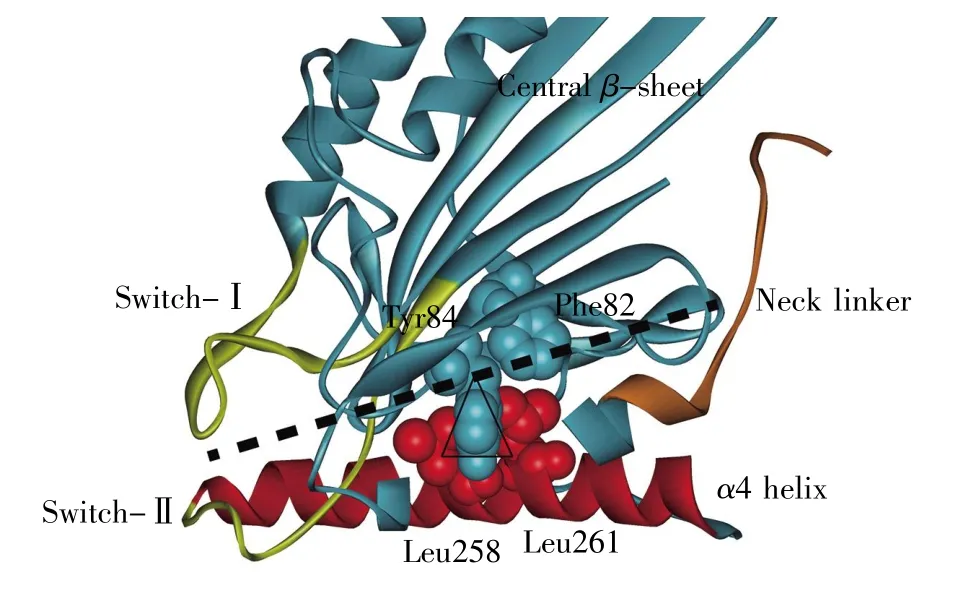
图4 驱动蛋白力学传递的“跷跷板”模型驱动蛋白中心β片用蓝色表示。Switch-Ⅰ和switch-Ⅱ组成了ATP结合腔,用黄色表示。α4螺旋用红色表示。图中颈链 (棕色)处于颈链结合腔。中心β片上的Tyr84和Phe82(蓝色CPK模式)以及α4上的Leu258和Leu261(红色CPK模式)组成了“跷跷板”的支点。黑色虚线是“跷跷板”板面示意图。Fig.4 “Seesaw”model of kinesin’s mechanical force transmissionThe central β-sheet of kinesin is colored in blue.Switch-Ⅰand switch-Ⅱof the ATP-binding pocket are colored in yellow and α4 helix is colored in red.The neck linker(brown)in this figure is in the neck-linker-docking pocket.Tyr84 and Phe82(shown in blue CPK mode)of central β-sheet together with Leu258 and Leu261(shown in red CPK mode)of α4 form the pivot of“seesaw”.The black dotted line represents the board of the “seesaw”.
3 马达结构域转动与颈链对接的关系
对于颈链向马达结构域对接的启动机制和对接过程,我们在综述文献[39]中进行了详细的讨论,此处不再赘述。值得注意的是,我们提出颈链在对接的初态构象中与β0保持了4个氢键相互作用。此相互作用是驱动蛋白马达结构域中心β片转动效果传递到颈链并启动颈链对接的关键。颈链的N端连接的是α6螺旋。中心β片的弯曲和转动效果无法直接影响α6螺旋和颈链。而β0直接与中心β片的β1相连,中心β片的转动直接带动β0的位移。此位移通过β0和颈链之间的4个骨架氢键传递到颈链,从而启动颈链向马达结构域的对接。我们的全原子和粗粒化分子动力学模拟结果也对此进行了证明[38,40~42]。
4 总结与展望
通过上述分析可知,驱动蛋白以两种方式利用ATP的能量。首先,驱动蛋白的ATP结合位点与高度带负电荷的ATP分子以及和ATP分子螯合的Mg2+发生特异性的结合,使得总能量降低,
由此产生的力使马达结构域的ATP结合位点产生一系列构象变化[9]。这些构象变化经过马达结构域其他元件的传递和放大,进而启动驱动蛋白的力产生过程。此后,驱动蛋白会催化ATP分子发生水解反应[23]。ATP水解反应是一个放能过程,
所释放的能量使得水解产物磷酸基团具有较高的能量,从而逃离驱动蛋白ATP结合位点周围氨基酸残基的束缚。在磷酸基团的释放过程中,会诱导ATP结合位点产生一系列的构象变化。这些构象变化减弱了马达结构域与微管的相互作用。当此头部与其下一个微管结合位点结合时,ADP分子释放,使驱动蛋白恢复到ATP结合之前的构象,从而使驱动蛋白的下一个循环得以进行[27~28]。分子马达研究的核心问题是其能量的利用和转化的机制问题。此问题的解决是我们彻底理解分子马达的标志。通过相关学者30多年的努力,
对于驱动蛋白能量转化问题的研究有了非常大的进展和突破。但是,我们还不能完全从氨基酸水平将整个过程描述清楚,需要进行更深入的研究工作。另外,由于核苷酸酶在核苷酸结合位点具有高度保守性,因此驱动蛋白能量转化机制问题的研究对于其他核苷酸酶结构-功能关系的理解也有非常大的指导意义。
