养分添加对白桦叶片气孔和气体交换异质性影响研究
2019-04-08刘力铭孙志虎李开隆龚丽芳
刘力铭,孙志虎,李开隆,龚丽芳
(东北林业大学 林学院,黑龙江 哈尔滨 150040)
气孔作为植物气体交换的主要通道,其特征由遗传因素与非遗传因素共同影响,多数植物气孔特征对环境因素变化较敏感,性状不稳定[1]。近些年来,因其具有指示大气CO2与气候变化的特点,从而使得利用化石气孔数据重建古气候成为可能[2-3]。气孔密度(SD)的变化能够体现植物耐旱性或营养摄取能力的调整[4]。随着叶片气孔特征与气体交换研究的不断深入,研究发现同一叶片不同区域(叶尖、叶中、叶基)气孔分布与气体交换具有差异[5-8],然而这种差异的发生仍无定论,普遍认为基因决定着气孔在叶片上的分布,然而外界环境差异也是植物生长过程中不可忽略的重要影响因素。研究结果表明,养分变化对叶片光合能力与SD均会产生显著影响[9-11]。林木光合特性方面的现有研究多集中于生态因子(如温度、CO2浓度、光照等)对整张叶片光合特征的影响方面[12-14],缺少同一叶片不同区域间光合和气孔特征的差异性研究。气孔作为气体交换的主要通道,在不同养分条件下叶片气体交换特征存有差异的同时叶片气孔分布特征(如SD)是否也发生相应的变化?同一叶片不同区域间的二者关系是否受到土壤养分状态的影响等问题研究却少见报到。
白桦Betula platyplylla是东北森林的重要组成树种,进行不同养分状态下白桦叶片气体交换特征研究对于揭示天然白桦在叶片水平适应环境变化的策略具有重要意义。本研究利用养分添加后白桦叶片不同区域气体交换特征与SD的调查结果,分析二者间的关系,评价养分差异和气孔密度变化对叶片气体交换特征的影响,以期为不同生境下白桦的适应机制研究和育林改造等营林技术提供理论依据。
1 材料和方法
1.1 试验材料
试验于东北林业大学强化育种基地6号温棚中进行。所选种子采自同一株母树。4月23日,将事先浸泡催芽处理过的白桦种子点播于25孔(每孔规格8 cm×8 cm×8 cm)的蜂巢状育苗盘(盘中育苗基质为草炭、沙子、黑土按照5∶2∶1的比例混合,未施肥前的土壤N、P、K、硝态N、氨态N、有效P、速效K含量分别为5.55、0.59、4.54 g/kg和82.36、13.13、10.70、92.95 mg/kg[10]),每孔点播种子3~4粒,共25盘。5月上旬,每孔定植1株白桦。6月23日,幼苗高度为10 cm左右时,将25盘的白桦幼苗分成25组(每组1盘,每组25株幼苗),按照L25(56)正交试验设计进行定量施肥(表1)。一次性增施N、P、K肥的最大量是按照连续72 d,每2 d浇灌1次,累计浇灌4 320 ml Hogland营养液的标准换算为化肥量一次性施入土壤,养分添加时所使用的N、P、K为分析纯级的过磷酸钙(P)、硫酸钾(K)和尿素(N)自行配置。养分增施时采用撒施方式添加P,采用水溶液的方式喷施N和K于土壤中。
1.2 试验方法
8月7—10日,每种处理均随机选择3株苗高为15 cm以上的幼苗为对象,选取施肥后新长成的苗茎的中上部位的完全展开叶,利用 Li-6400 便携式光合测定系统测定叶片3个区域(叶尖、叶中和叶基)的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs),测量时确保叶片3个区域在叶室中没有重叠。完后,将相应叶片摘下进行3个区域SD的调查。SD调查时,3个区域均选择3个视野(共9个视野点,面积为0.3 mm2)进行拍照,所用拍照设备为基恩士(KEYENCE)超景深三维数码显微系统VHX-5000。利用Image-Pro plus6.0(Media Cybernetics公司)软件计量每个视野中的气孔数,并计算气孔密度(个/mm2)。
1.3 统计分析
利用Excel 2010对数据进行描述统计分析;利用 SPSS19.0(SPSS Inc.,Chicago,IL,USA)进行正交试验数据分析;利用Duncan法进行不同施肥水平间的差异比较;并对气体交换各项参数与气孔密度进行Pearson相关性分析。

表1 L25(56)正交试验设计(每株苗各类化肥施肥量)†Table1 L25(56) orthogonal experiment design (the amount of different fertilizers for per plant)
2 结果与分析
2.1 不同施肥水平对不同区域气体交换与气孔密度的影响
P、N因素对白桦叶片叶尖Pn影响均显著(P<0.05)。虽然K因素对白桦叶尖Pn影响不显著但交互作用对叶尖Pn影响均极显著(P<0.01)。N因素下N2水平的Pn的较高,然后随着N施入量的减少逐渐降低,说明N1的施入量过高不利于叶尖的光合作用。P因素下P4水平的Pn较高,随着P施入量的减少叶尖的Pn呈现出先增加后降低的趋势,说明少量的P会增加叶尖的Pn。N、P、K以及交互作用对白桦叶片叶尖的Tr影响均不显著。N、P、K各个因素对叶尖Gs影响均不显著,而NP、PK、NPK交互作用对Gs影响显著(P<0.05)。N、P、K以及交互作用对白桦叶片叶尖SD影响均不显著。综上,N、P对叶尖Pn的影响较大,单一的N、P、K养分变化对白桦叶片叶尖的Gs影响并不大。而交互施肥导致的养分变化则会强烈影响叶尖的Gs。N、P、K及其交互作用对白桦叶片叶尖的Tr与SD影响均不显著(表2)。
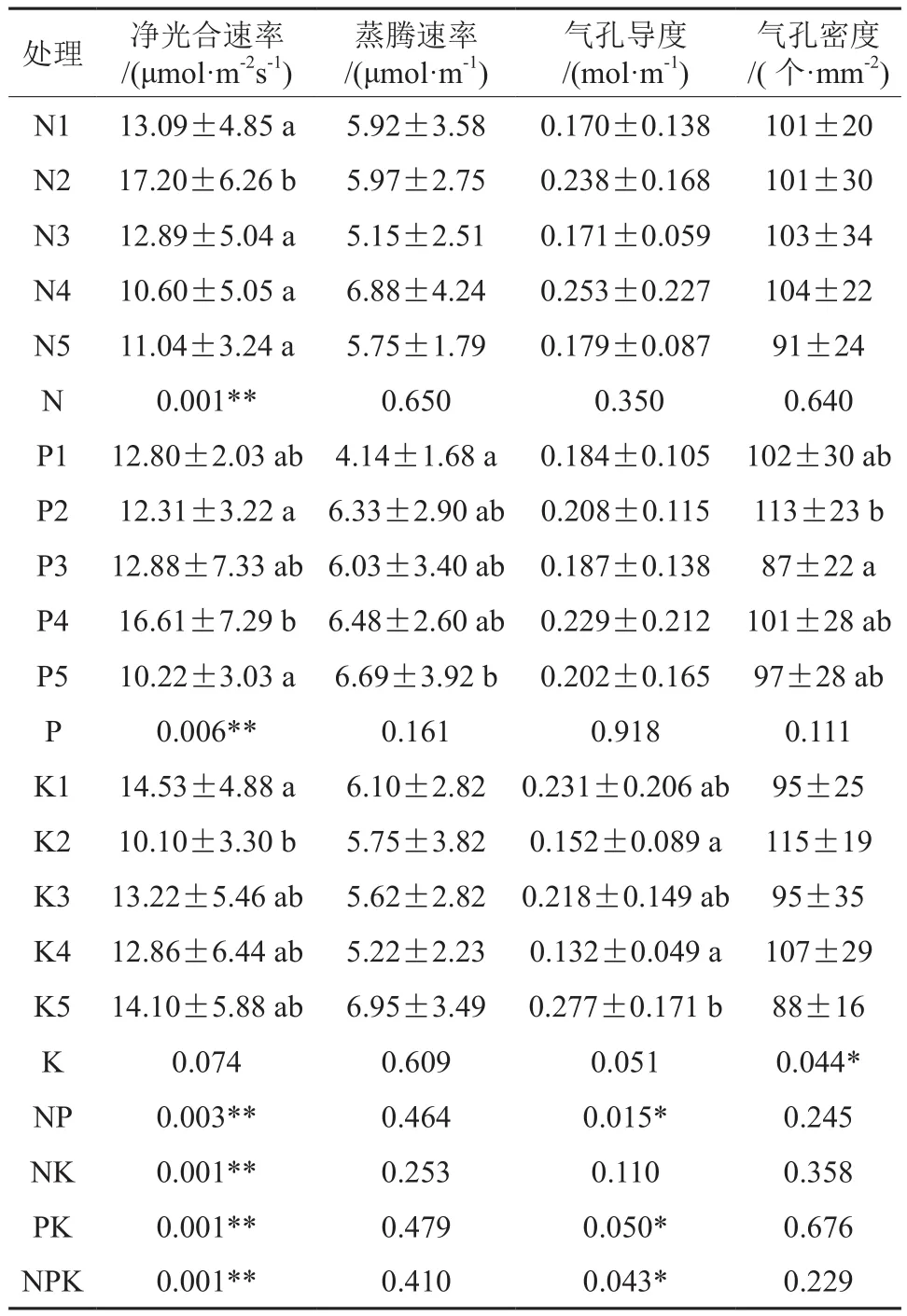
表2 N、K、P及其交互作用对白桦叶尖气体交换与气孔密度的影响†Table2 Effects of N, K, P and their interactions on gas exchange and stomatal density of leaf tip
N、K、P及其交互作用对白桦叶中气体交换与气孔密度的影响见表3。N、P因素对叶中Pn影响均显著(P<0.05),而K因素对叶中的Pn影响不显著,但交互作用对叶中Pn影响均极显著(P<0.01),这与其对叶尖的影响一致,叶中Pn数值远远小于叶尖,并且叶中的Pn与叶尖Pn的变化较为相似。与叶尖相同各因素对白桦叶片叶中的Tr影响均不显著。N、P、K对叶中Gs影响均不显著,而交互作用对叶中Gs影响均显著(P<0.05)。虽然NP、NK、PK交互作用对白桦叶中SD影响均不显著,但是与叶尖不同的是P不同水平对SD影响显著(P<0.05),K不同水平对SD影响极显著(P<0.01),并且N、P、K交互作用对白桦叶中SD影响显著(P<0.05)。综上,施肥对叶中的SD影响与叶尖较不同,并且气体交换参数均小于叶尖,除此之外施肥对叶中与叶尖的影响大致相同。

表3 N、K、P及其交互作用对白桦叶中气体交换与气孔密度的影响†Table3 Effects of N, K, P and their interactions on gas exchange and stomatal density of leaf middle part
各个因素水平对白桦叶基Pn影响与叶尖、叶中相同。但与叶尖和叶中不同是P对叶基的Tr影响显著(P<0.05),其他各因素水平对白桦叶片叶基的Tr影响均不显著。N、P、K对叶基Gs影响均不显著,而NP、PK交互作用对Gs影响依然显著(P<0.05)。这一结果与叶尖相同。N、P、K以及NP、NK、PK交互作用对白桦叶片叶基SD影响均不显著(P<0.05),这与叶尖的结果一致(表4)。
通过观察不同处理下叶尖、叶中、叶基的气体交换与SD发现不同处理叶中与叶基Pn、Tr、Gs的结果具有相同趋势并且大小相接近,但叶中略高于叶基,虽然叶尖总体趋势与叶中、叶基相同,但叶尖的Pn、Tr、Gs明显大于叶中与叶基的Pn、Tr、Gs。因此白桦叶片气体交换异质性主要存在于叶尖对叶中和叶基的异质性上。而SD在叶片不同区域分布较均匀,但叶中SD受养分变化要比叶尖、叶中明显,并且相比Tr、Gs与SD,白桦叶片各个区域Pn受养分变化影响较大(表2—4)。

表4 N、K、P及其交互作用对白桦叶基气体交换与气孔密度的影响†Table4 Effects of N, K, P and their interactions on gas exchange and stomatal density of leaf base
2.2 白桦叶片不同区域净光合速率与蒸腾速率、气孔导度、气孔密度的相关关系
白桦叶片各区域的Pn和Tr、Gs、SD的相关性分析见表5。叶中与叶基的Pn和Tr显著。而叶尖与叶中、叶基不同,叶尖的Pn和Tr相关性不显著。因此白桦叶片不同区域的气体交换异质性也表现在Pn和Tr的异质性上。Pn与Gs在叶片各个区域均有极其显著的正相关。Pn与SD在叶片各个区域均没有显著的相关性。此外不同区域气体交换数值与SD相关性均不显著。综上说明白桦叶片不同区域气体交换特征存在差异可能是由于气孔密度以外的因素引起。

表5 白桦叶片不同区域净光合速率与蒸腾速率、气孔导度、气孔密度相关关系†Table5 Correlation between net photosynthetic rate and transpiration rate, stomatal conductance and stomatal density in different regions of B.platyphylla leaf
3 结论与讨论
N、P、K是植物成长过程中最重要的三大元素,适量的施入N、P、K会有利于叶片光合作用,而过多的施入不仅不利于叶片光合有机物积累,相反还会起到抑制作用。本试验通过正交试验的优势来达到养分条件的不同变化,从而观察叶片气体交换与气孔密度在叶片水平上的变化。其目的在于更好地了解环境条件变化的同时,植物器官对其反应的方式与策略,其结果可以为后续的科学问题提供假设与猜想。本研究表明白桦幼苗叶片光合作用受养分变化影响较大,不论叶尖、叶中、叶基都达到了极显著。尤其是N和P对其的影响要比K明显。研究表明增施N肥能有效地提高叶片中叶绿素含量以及Rubisco酶含量,从而提高叶片的Pn,而过量的施入会导致Rubisco酶含量超出正常值,降低叶片的光合速率[16]。本研究中叶片各个部位由于N1水平N施入量的过高导致Pn下降,N2水平的Pn最高,随后随着N含量的降低逐渐减小。P作为光合磷酸化的主要原料对光合作用也同样具有重要影响。本研究中叶片各个部位随着P肥施用量的降低呈现先减少后增加的趋势,并在P4水平下最高。说明过高的P也会抑制叶片的Pn。此外本研究发现尽管不同养分条件对光合作用影响较大,但是对叶片Tr影响不大,仅发现P对叶基Tr有显著影响。因此说明在养分条件改变时蒸腾作用的强度不一定会与光合同步。养分条件的改变并不会像其他因素改变光合作用强度的同时使得蒸腾也随之发生很大的变化,例如温度、有效光辐射、水分、CO2浓度等。本实验中蒸腾没有像光合那样受控于养分条件,分析其原因可能与水分限制有关。本研究还发现不同养分条件下Gs也有显著变化,虽然N、P、K单个营养元素对其影响不显著,但交互作用显著影响白桦叶片气孔导度。而多数研究表明Gs与Tr均有很强的相关性。例如郑凤英[17]的研究表明在水分胁迫下Gs与Tr呈正相关。但王春枝[19]通过施肥的角度研究表明Gs与Pn是一致的[18]。并有研究表明在未受到水分亏缺的情况下,Gs与Tr呈正相关,但是从对Gs的影响大小看,与光合作用的影响一致。因此分析其原因可能由于本实验中水分不充足,植物保水功能增强,由于其光合作用的改变使得进入叶片中的CO2量发生改变从而间接影响了白桦叶片Gs的显著变化,而并非由于蒸腾作用导致其改变。这也可以解释为什么多数研究中,施肥过后水分利用效率大幅增强的原因。
对叶片气孔分布的报道已有很多[20-21]。树木叶片在发育过程中,很大一部分原因是基因控制着细胞分裂与分化导致SD与气孔分布在叶片上的异质性。Poole等将气孔在叶片上的变化归结于3个原因,一是气孔和表皮细胞的不均匀分化,导致响应的细胞数量变化(分化假说),二是表皮细胞膨胀导致气孔间距不均匀(膨胀假说),三是前二个原因的混合(混合假说)[22]。然而环境因素也是影响气孔发育的一个重要因素,如,辐射强度、湿度、温度、大气CO2和臭氧含量,土壤水分和营养的可用性[23]。也有研究表明植物可通过已经形成的器官感受外界环境后,将环境信号反馈给植物本体影响即将形成的组织器官的形态,进而来适应环境变化[24-25]。而气孔因素对光合CO2同化的影响也一直存在争议,部分研究表明叶片气体交换与气孔的因素相关性很强[26],也有研究表明气体交换并非完全受控于气孔因素[27]。本研究中白桦SD在叶片不同部位没有显著差异,SD与各项气体交换之间未达到显著水平,说明白桦叶片气体交换异质性主要由SD以外的因素引起的。这与Nardini等(2008)的研究相似,其在烟草叶片气体交换的空间异质性调查中发现叶尖对水蒸气的传导速率比叶基高18%,叶尖导电率也比叶基部分高30%[28]。而叶尖与叶基的SD却没有明显差异,同样说明叶片各部位功能异质性与SD差异并非同步。对这种叶片光合异质性的看法也有不同的解释,Strain等[5]认为叶片从叶基部位开始生长,叶基部位要比叶尖部位的组织年轻,而较老的组织更能适应强光照,Xiong等[6]通过调查两个品种的水稻气体交换特性参数与叶片结构的化学特性表明气孔和叶绿素含量从叶基部到叶尖部分逐渐增加。Xiao等[7]的研究表明即使叶绿体位于叶子表面的距离相同,其受光环境也会有截然不同,由于叶片不同区域的叶表皮组织差异,也会影响叶片光合效率。也有研究表明N对提高光系统Ⅱ(photosystemⅡ,简称PSⅡ)有显著作用[29],盐胁迫与高温均会对PSⅡ的异质性产生影响[30-31]。由此表明叶片光合异质性可能受控于多种因素的共同影响。王碧霞等[8]利用网格与二维成图的方法对青杨雌雄叶片各区域气孔分布及气体交换特征的研究发现,不同区域气孔分布于气体交换有明显的不同,在雌株中叶基的Pn最低,叶尖的Pn最高,并相差3~4倍以上。虽然雌雄同一叶片的Pn都具有异质性,但异质程度却不相同,表明不同品种的植物光合异质性也存在强弱区别。而有时环境的变化甚至会导致光合异质性发生强烈变化。XU等[32]的研究表明在自然状态下雌雄异株的杨树,两性叶片Pn从叶尖到叶基逐渐下降。在盐胁迫下雌株却出现了相反的趋势,表明不同类型的植物应对不同环境变化采用的策略是不同的。本研究显示在不同施肥处理下白桦同一叶片叶中的Pn要比叶基的Pn高一些,不同施肥处理两者都具有较高的一致性。而除叶尖的Pn由于施肥处理后变化幅度较大并没有与叶中、叶基的一致程度高外,不同施肥处理下白桦同一叶片的叶尖、叶中、叶基的Pn并没有因为养分变化而发生差异规律的较大变化。叶尖的Pn均大于叶中与叶基以及叶中也略高于叶基的这种差异规律也没有因为施肥出现相反的趋势,表明白桦叶片光合速率在不同养分添加的情况下其应对方式变化不大,值得注意的是白桦叶片不同区域的光合与蒸腾间的关系也是不同的,说明叶片气体交换的异质性也存在于光合与蒸腾的相关关系上。虽然本研究可以表明白桦不同区域叶片各项气体交换之间存在差异性并且与叶片SD无关,但未能验证具体是什么原因导致以及哪种因素对其影响较大。分析白桦这种气体交换异质性的原因可能由于叶片不同区域的叶绿素含量、酶活性、养分吸收能力、以及内部结构与PSⅡ等的不同导致气体交换异质性的发生。因此在今后研究人员可以针对叶片不同区域的叶绿素含量、酶活性、养分吸收能力以及内部结构等进行开创性的实验设计与方法来验证叶片不同区域生理变化的原因。
