油茶果实生长高峰期养分分配特征
2019-04-08陈隆升陈永忠许彦明彭映赫
陈隆升,罗 佳,陈永忠,许彦明,张 震,唐 炜,彭映赫
(1.湖南省林业科学院,湖南 长沙 410004;2.国家油茶工程技术研究中心,湖南 长沙 410004)
油茶Camellia oleifera是一种生长相对较慢的长寿树种,生命周期长,从花芽分化、开花授粉、果实生长发育成熟整个过程历时约17个月。秋花秋实,往往果期尚未结束,花期又至,所以民间称之为“抱子怀胎”[1],这是油茶植物异于其他果树的一大特征。
3月中旬左右子房膨大初具果实雏形,油茶果实一般在4月下旬—6月中旬和7月上旬—8月下旬左右生长较快,有2个生长高峰[2-4]。此间的油茶果实生长发育伴随着花芽分化与春梢、夏梢、秋梢的抽梢生长;果实生长、花芽分化与抽梢生长都是紧密联系并相互影响的,营养生长和生殖生长相互交错。期间的养分充足与否不仅影响当年的产量,而且影响花芽分化,对第二年的产量也产生决定性的影响[1]。油茶果实快速生长的这段时间频发高温、干旱天气,如养分供应不足,常常造成大量落果、花芽发育不良、果实发育生长不良导致产量与品质受到严重影响。因此,充分掌握油茶果实快速生长期的油茶养分分配规律对于科学指导施肥,保证油茶丰产、稳产具有重要的作用。目前有不少关于油茶树体各器官营养元素的吸收、累积和分配规律等方面报道[5-7],但涉及营养元素、油茶器官的种类不够全面[8-12],虽对春稍生长期[10]、花期[13]、果实成熟期[14]等油茶关键物候期的养分分配规律进行研究,但对油茶果实生长高峰期这一最关键的生长期的养分分配规律研究未见报道。因此本研究通过研究油茶果实生长高峰期各器官的N、P、K、Ca、Mg等大量元素以及Cu、Fe、Zn、Mn等微量元素含量的分配规律,为油茶营养诊断、科学准确的配方施肥提供理论依据。
1 试验材料与方法
1.1 试验地概况
试验样地位于湖南省常宁市荫田镇白马村,为丘陵山地,112 °37 ′4″E,26°20′40″N,海拔 82 m。属亚热带季风性湿润气候,年均温 18.1 ℃,年均降水量1 436 mm。基地现有油茶林350 hm2,于2010年3月采用两年生裸根苗造林,品种主要为湘林系列良种。土壤母质为第四纪红壤土壤贫瘠,土壤为红壤,具体土壤理化性质,见表1。

表1 试验油茶林土壤( 0~60 cm) 的养分特征Table1 Nutrient characteristics in arable soil layer (0~60 cm) of C.oleifera test forest
1.2 试验设计
选择具有典型代表性、林分生长、郁闭度、立地条件基本一致的3个样地,样地面积为20 m×20 m,样地四角处竖 PVC管标志。在果实成熟期(2015年10月16日)通过对果实、树型的观测,每个样地选择果实性状、树型一致的植株5株做为定点观测标准样株。
1.3 样品采集
植物样品采集:分别于2016年5月16日(代表果实第一次生长高峰期)、2016年8月10日(代表果实第二次生长高峰期)对每个样地内的5 株标准样株进行采样。采集叶片为树冠层中部东西南北方向生长状况相对一致的1年生枝条从基部往上第 3 ~ 5 片成熟叶;在采集叶片的同时采集该1年生枝条;采集果实为分枝层中部东西南北方向成长情况基本相同的树冠外围果实。采集根时在每株标准株基部挖取南北方向、深40 cm剖面,收集直径<2 mm的细根,并根据细根的颜色、弹性、外形挑出活细根,装入纸袋;为了减少取样对树体影响,采集树干时在样株上部(距离顶端10~15 cm)钻取直径0.5 cm的孔进行取样。取每个样地采集的叶、果、根、枝和干分别混匀后,每个样地为1个重复,共3 个重复,每个重复鲜质量约为300 g(干与根系为30 g),于实验室 65 ℃烘箱中,烘约48 h,直至恒质量,样品粉碎过 0.25 mm 筛后测定养分元素含量。
1.4 样品的测定分析
全 N、P、K 含量用H2SO4-H2O2消煮后,分别用凯氏定氮法、钼锑抗比色法和原子吸收法测定,Cu、Fe、Zn、Mn(均为全量)用HP3510原子吸收分光光度计测定[15]。测定结果以单位质量的养分含量表示, N、P 与P、K化学计量比采用质量比表示。
1.5 数据分析
采用SAS6.0和Excel进行数据处理与统计分析,采用SAS6.0中析因设计的方差分析对油茶不同时间(5月、8月)、器官(果、叶、枝、干、根)的营养元素进行分析。
2 结果与分析
2.1 不同月份与器官油茶大量元素变化
方差分析表明,油茶N、P、K、Ca、Mg等元素含量均受不同器官和不同时间的影响,并呈现极显著(P<0.01)的差异(表2)。油茶各器官的N含量均以叶片最高,5月与8月分别为26.77 g·kg-1、20.61 g·kg-1。8 月油茶果、叶、枝与干的N含量比5月分别下降了36.70%、23.01%、32.60%、9.71%,而根系的N量则相反,增加了4.97%。
8月油茶各器官P含量均小于5月, 果、叶、枝、干、根分别下降了42.00%、27.77%、28.30%、2.94%、41.67%。各器官中P的含量以叶片最高,枝次之,但叶与枝的P含量差异很小,叶片5月与8月的P含量分别为0.54、0.39 g·kg-1,枝则分别为0.53、0.38 kg-1。5月各器官的K含量均高于8月。5月各器官K的含量以叶片最高,为7.83 g·kg-1,8月则以果实含量最高,仍维持在较高的水平为6.32 g·kg-1,仅比5月下降了2.91%,而其它器官则下降幅度为 42.02%~59.58%。
各器官Ca的含量以叶片最高,5月与8月分 别 为 7.61、11.01 g·kg-1。5月 果、叶 与 枝 的Ca的含量低于8月,8月分别增加了90.60%、44.68%、43.48%,而干与根则相反,分别下降了2.08%、16.67%。各器官Mg的含量以根最高,5月 与 8月 分 别 为 1.79、1.29 g·kg-1;5月 果、叶与根的Mg的含量均高于8月,分别下降了39.06%、34.44%、27.93%。枝与干则相反,但变化较小,仅增加了3.19%、6.58%。
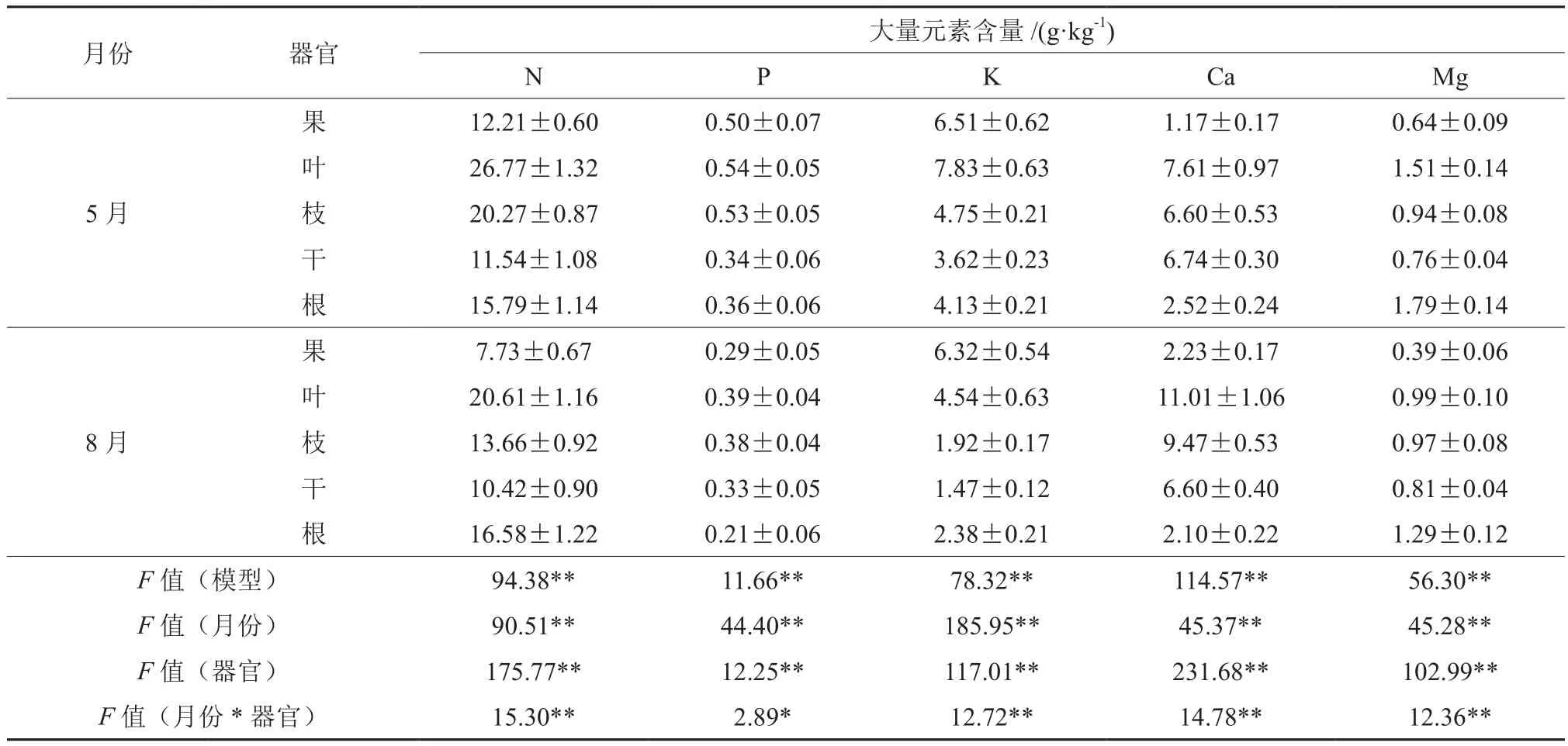
表2 油茶各器官大量元素含量Table2 Macro-element contents of C.oleifera organs
2.2 不同月份与器官油茶叶微量元素变化
方差分析表明,油茶Cu、Fe、Zn、Mn等微量元素含量均受不同器官和不同时间的影响,并呈现极显著(P<0.01)的差异(表3)。5月各器官Cu含量均显著高于8月,8月果、叶、枝、干、根分别下降了17.34%、33.91%、14.51%、22.97%、11.41%。5月与8月油茶各器官的Cu含量为枝>干>叶>果>根,5月枝Cu含量最高达到了 18.33 mg·kg-1。
5月果、枝、干、根Fe含量显著高于8月,分别下降了73.17%、39.06%、88.98%、43.61%;而叶则相反,增加了19.20%。5月Fe含量为根>枝>干>果>叶, 8月Fe含量为根>枝>叶>果>干,其中5月根的Fe含量最高,达到了748.85 mg·kg-1,8月干Fe含量最低为54.16 mg·kg-1。
5月果、叶Zn含量高于8月,8月分别下降了33.51%、10.44%;枝、干、根则相反,分别增加了6.53%、9.85%、58.07%。5月与8月Zn含量为枝>干>根>叶>果,8月枝Zn含量最高达到了50.76 mg·kg-1,8月果实中Zn含量最低,仅为6.33 mg·kg-1。
5月各器官Mn含量均显著低于8月,8月分别增加1.36%~171.15%。5月Mn含量为叶>枝>干>根>果,8月Mn含量为叶>枝>根>干>果,以8月叶Mn含量最高,达到了1 159.05 mg·kg-1,5月果Mn含量最低,仅为211.02 mg·kg-1。
2.3 油茶叶养分的化学计量比
5月份油茶叶片N∶P与P∶K值分别为50.26、0.07,8月份叶片N∶P与P∶K值分别为53.12、0.09,分别增加了5.69%、28.57%。N∶P高于16,表明油茶幼林主要受P的限制,与5月相比,8月P的限制进一步加剧(表4)。
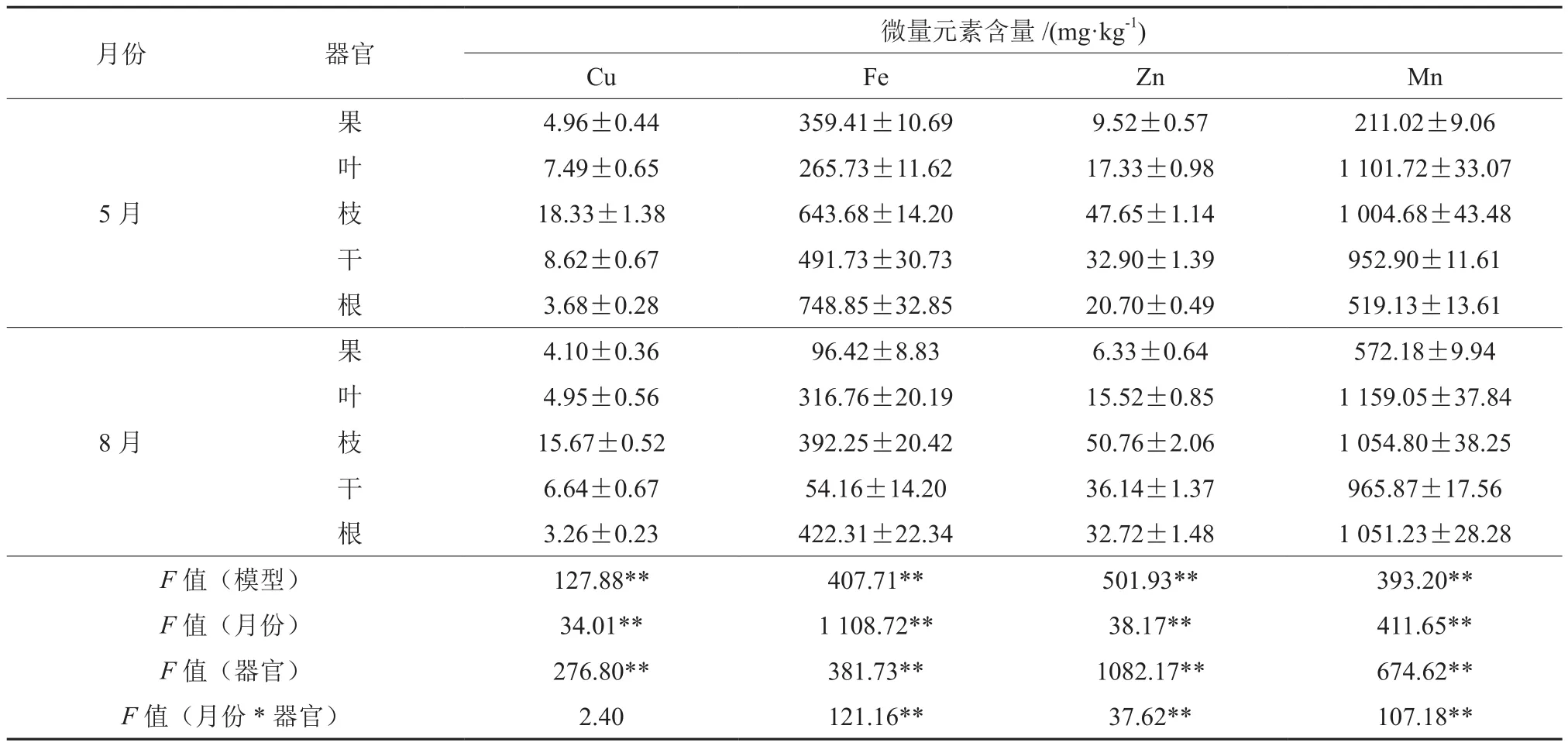
表3 油茶各器官微量元素含量Table3 Trace element contents of C.oleifera organs
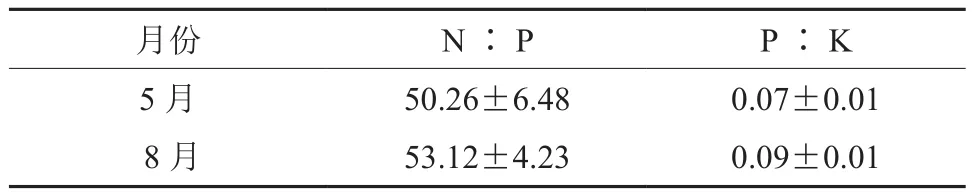
表4 叶养分元素化学计量特征Table4 Stoichiometric characteristics of nutrient elements of C.oleifera leaves
3 讨论与结论
4月下旬—6月中旬和7月上旬—8月下旬为油茶果实2个生长高峰,8月下旬之后油茶果实大小基本稳定,并积累大量糖类物质,进入油脂合成积累高峰期[16-17]。同时笔者观测本试验林发现5月为春梢成熟期,5月下旬,在当年成熟的春梢上开始分化出大量花芽,6月中旬花芽基本形成,夏梢开始生长。油茶果实生长高峰期同时又是花芽生长发育、新梢生长关键期,因此,油茶树体需要大量N、P、K养分,特别是油茶果实快速生长与花芽的分化。本试验中除根系N含量小幅度增加之外,8月其余各器官的N、P、K浓度均低于5月,造成这种现象可能有3个方面的原因,一是土壤养分供给不足,与其他母岩发育的油茶林土壤相比[18-19],本试验林地土壤有效N、P、K明显偏低,尤其是有效P、K含量(表1)。二是枝、叶、干、根自身生长稀释了养分的浓度,尤其是果实的快速生长与花芽的生长发育;三是相对应叶等“源”器官,果实与花芽等“库”器官对养分的分配具有更大的“吸引力”[20],在树体整体养分不足的情况下,大量的 N、P、K等元素向果实与花芽转移,以满足其生长发育的要求,导致了叶、枝、干等其它器官养分含量降低。
8月各器官K含量以果实最高为6.32 g·kg-1,仅比5月下降了2.91%,而其它器官则下降幅度为42.02%~59.58%。这主要是一方面油茶果实快速的生长从枝、叶、干、根等其他器官吸收了大量的K元素,另一面是积累充足的K元素,有利于促进糖分的转化与运输,使得果实快速积累大量的光合产物[21],为之后的油脂转化与果实成熟积累充分的营养,核桃上也有类似的研究结果[22]。
N∶P高于16表明植物受P的限制[23-26],本试验中油茶5月、8月油茶叶片N∶P 分别为50.26、53.12,说明本试验油茶林主要受P的限制,且8月P的限制进一步加剧。与5月相比,8月油茶叶叶片P、K含量显著降低,但油茶叶片中K的下降幅度要高于P 导致了P∶K增加,这主要是由于8月正处于油茶油脂合成关键期,叶片中大量的K被转移利用以促进油脂的合成,这也是油茶果实在8月仍可以维持较高K含量的重要原因之一。因此,对挂果的油茶林在提供充足的N的同时,在果实快速生长期及时补充P、K元素至关重要。
Ca主要存在于叶与老的器官、组织中,一般很难移动,钙离子(Ca2+)是一种重要的第二信使,参与调节植物的生长发育和对环境的适应[27-28]。本试验发现,油茶吸收Ca含量为叶>枝>干>根、果,说明叶片作为光合作用与各类逆境反应的重要器官,积累了大量钙离子(Ca2+)以满足生长发育和适应环境的需要。与5月相比,8月叶、果Ca含量增加了44.68%、90.60%,这可能是8月处于季节性高温干旱期,同时也是油茶果实油脂转化合成关键期,果实与叶片吸收积累更多的钙离子(Ca2+)以满足各类生理活动所需。
与5月相比,8月油茶 果、叶、根Mg含量大幅度下降,枝与干则小幅度上升,可能是由于随着果实快速生长与花芽分化,从土壤吸收以及原本存储在叶、根的部分Mg元素优先转移到了果实与花芽,在满足果实与花芽生长的需要之后储存了一部分在枝与干,导致了枝与干Mg含量的小幅度上升。果实的Mg含量下降则是其果实快速膨大导致Mg浓度降低的结果。
Cu、Fe、Zn、Mn 等4种微量元素中,Zn为可转移再利用的元素,且是叶绿素合成的必须元素[27],8月果实与叶片Zn分别下降了33.51%、10.44%,这可能8月叶片已完全成熟,叶片中的部分Zn转移到花芽或枝、干、根中,而8月果实Zn含量下降则可能是其快速生长导致的Zn浓度被稀释造成的。与5月相比,8月叶片的Fe、Mn含量增加了19.20%、5.20%,表明随着时间的延长更多的Fe、Mn被固定、储存于叶片中,参与光合、氮代谢等过程[29]。
