千里光茎和叶的差异表达基因分析
2019-04-02钱秋博言贺莉芳杨春先左青青
谢 欣, 钱秋博言, 贺莉芳, 李 林, 杨春先, 左青青, 钱 刚,①
(1. 遵义医科大学细胞生物学教研室, 贵州 遵义 563000; 2. 黔南民族医学高等专科学校护理系, 贵州 都匀 558003)
随着高通量测序(high-throughput sequencing)技术的快速发展,用于数字基因表达谱(digital gene expression profiling, DGE)研究的深度测序技术不但广泛用于有参考基因组序列的物种分析,而且能够直观显示基因组的功能元件和剪切方式,确定基因的转录结构,量化各转录本在发育过程中和不同条件下表达水平的变化,以及对所有转录产物进行分类[1],从而实现对无参考基因组序列的物种进行转录组学研究[2-3]。由于转录组测序蕴含了大量的基因表达信息和数据量,能够更加精确地定量分析特异基因的表达水平和等位基因转录本的特异表达,目前转录组测序已成为了解控制数量性状的基因组大规模表达情况的重要手段[4]。近年来,国内外研究者对部分经济作物和模式植物[5-8]进行了转录组测序研究,但在中医药领域研究中却处于起步阶段。
千里光(SenecioscandensBuch.-Ham. ex D. Don)隶属于菊科(Asteraceae)千里光属(SenecioLinn.),为重要的传统抗菌中草药,主要用于治疗风热感冒、目赤肿痛、泄泻痢疾及皮肤湿疹疮疖等[4,9],具有良好的开发和利用前景。目前,关于千里光的研究主要集中在化学组分和药理学作用机制等[9]方面,而关于其功能基因组学的研究却极其匮乏。相关研究表明:千里光不同品种的抗菌性差异显著[4],其不同部位的药效也有明显差异[10],因此,探明千里光不同器官的功能基因差异表达对于了解其药用成分积累和主要药理作用等具有重要意义。
鉴于此,作者采用高通量测序技术对千里光强抗菌性植株SC-32茎和叶的cDNA文库进行转录组测序分析,筛选出差异表达基因,并对这些差异表达基因进行功能分析,以期揭示千里光茎和叶的差异表达基因,为采用比较基因组学(comparative genetics)方法筛选菊科近缘药用植物茎和叶器官分化基因及其功能验证提供参考。
1 材料和方法
1.1 材料
以移植到遵义医科大学细胞生物学教研室中药材种质资源大棚内的野生千里光(来自贵州省遵义市板山地区)强抗菌性植株SC-32为实验材料,经遵义医科大学细胞生物学教研室钱刚教授鉴定。
1.2 方法
1.2.1 RNA提取和cDNA文库构建 取千里光1年生植株从上到下第7枚叶以上部位的茎和叶样品各3份,每份0.1 g,置于液氮中充分研磨;采用Trizol试剂盒〔天根生化科技(北京)有限公司〕提取总RNA;用质量体积分数1%的琼脂糖凝胶和NanoDrop 2000微量分光光度计(美国NanoDrop公司)检测提取的总RNA的纯度、浓度和完整性。
取检测合格的总RNA约1 μg,用带有Oligo(dT)的磁珠富集mRNA;加入5× fragmentation buffer,随机打断mRNA;以mRNA为模板,采用GeneRacer试剂盒(美国Invitrogen公司)构建cDNA文库。
1.2.2 转录组测序及数据分析 使用Illumina HiSeqTM2500高通量测序平台(美国Illumina公司)对cDNA文库进行转录组测序。利用Trinity软件将Illumina HiSeqTM2500高通量测序平台得到的raw data按顺序拼接成contig,利用TGICL软件进一步处理得到unigenes。利用FASTQC软件对测序数据进行质量控制,剔除只含有测序接头序列的reads、含N比例大于10%的reads以及低质量reads(Q≤10的碱基数占整条read碱基数的50%以上),从而获得高质量clean data;使用SOAPaligner/SOAP2软件将clean reads比对到青蒿(ArtemisiacarvifoliaBuch.-Ham. ex Roxb.)的参考基因组中,每个clean read最多错配5个碱基,统计clean reads在参考基因组中的覆盖度;采用FPKM(fragment per kilobase of exon model per million mapped fragment)法计算基因表达量,并以FC≥2且FDR<0.01为筛选标准(FC为差异倍数,FDR为错误发现率),利用DEGseq软件[11]筛选差异表达基因;利用BLAST软件(http:∥blast.ncbi.nlm.nih.gov/Blast.cgi)将unigenes序列比对到NCBI的Nr、Swiss-Prot、KEGG和COG(E值小于1×10-5)数据库中,利用Blast2GO软件根据Nr注释信息进行GO注释,并利用WEGO软件[12]对全部unigenes进行GO功能分类统计。
2 结果和分析
2.1 转录组测序结果
采用Illumina HiSeqTM2500高通量测序平台对构建的千里光茎和叶的cDNA文库进行转录组测序,在对测序数据进行质量控制后,将clean reads与青蒿参考基因组进行比对。结果显示:从千里光茎和叶的cDNA文库中分别获得26 147 016和26 996 119个clean reads,Q30值分别为95.17%和94.74%。在千里光茎和叶的cDNA文库中,比对到青蒿参考基因组中的clean reads(mapped reads)分别有20 722 491和21 610 120个,所占比例分别为79.25%和80.05%。其中,比对到参考基因组惟一位置的clean reads(unique mapped reads)分别有2 674 440和2 516 239个,所占比例分别为12.91%和11.64%;而比对到参考基因组多个位置的clean reads (multimapped reads)分别有18 048 051和19 093 881个,所占比例分别为87.09%和88.36%。
2.2 差异表达基因筛选结果
差异表达基因筛选结果(图1)表明:从千里光茎和叶中共筛选到10 991个差异表达基因,以茎中差异表达基因的表达量为准,叶中上调的差异表达基因有5 542个,占茎和叶差异表达基因总数的50.42%,而叶中下调的差异表达基因有5 449个,占茎和叶差异表达基因总数的49.58%。在筛选差异表达基因过程中,还发现41 045个表达量无显著差异的基因,并且,多数差异表达基因集中在-5≤log2(FC)≤5、-log10(FDR)≤25区域。
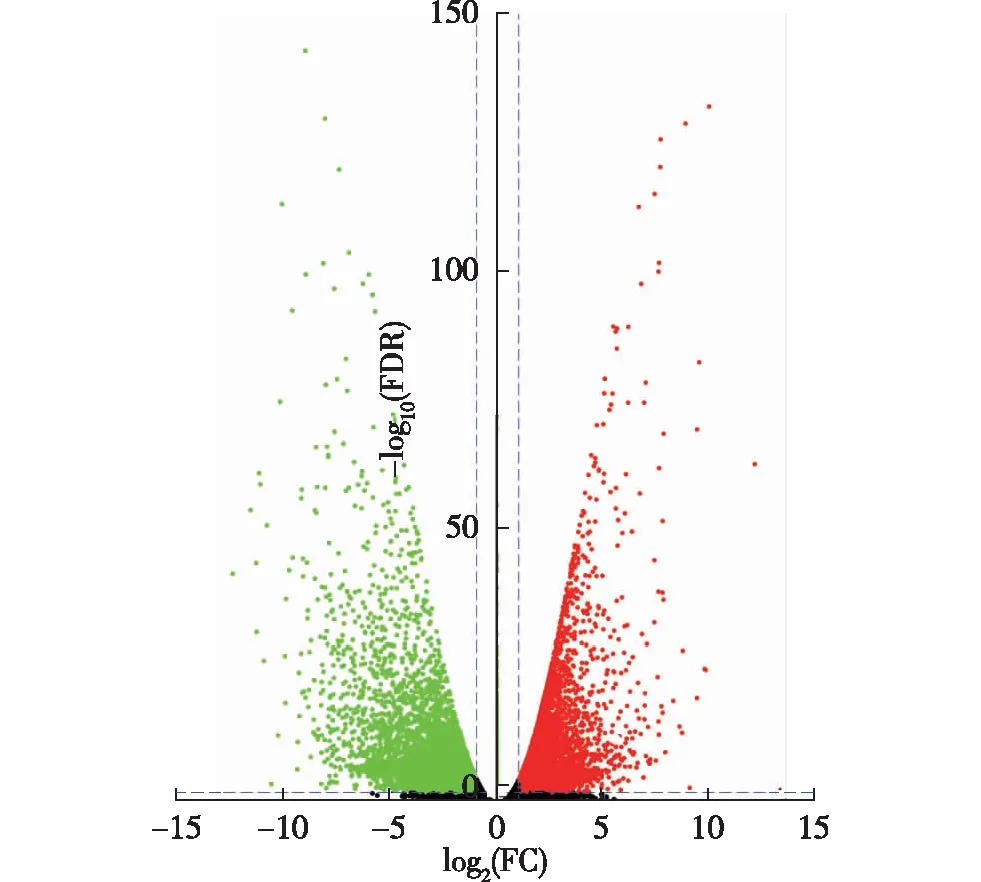
FDR: 错误发现率 False discovery rate; FC: 差异倍数 Fold change. 红点表示上调的差异表达基因 Red dots represent up-regulated differentially expressed genes; 绿点表示下调的差异表达基因 Green dots represent down-regulated differentially expressed genes; 横向虚线以下及纵向虚线间的黑点表示表达量无显著差异的基因 Black dots below horizontal dashed line and between vertical dashed lines represent genes without significant difference in expression level.
2.3 差异表达基因功能分析
2.3.1 GO功能分类分析 采用BLAST软件对千里光茎和叶中获得的clean reads进行比对,共有61 171个unigenes被注释,其中差异表达基因有7 683个。GO功能分类结果表明:这些差异表达基因的功能被分成生物过程(biological process)、细胞组分(cellular component)和分子功能(molecular function)3个大类,并被细分成53个亚类(表1)。在生物过程大类中,被注释为代谢过程(metabolic process)、细胞过程(cellular process)和单一生物过程(single-organism process)的全部unigenes和差异表达基因数量明显高于其他生物过程;在细胞组分大类中,被注释为细胞(cell)、细胞成分(cell part)、细胞器(organelle)、细胞膜(membrane)、细胞器成分(organelle part)和细胞膜成分(membrane part)的全部unigenes和差异表达基因数量亦较高;在分子功能大类中,被注释为催化活性(catalytic activity)和结合(binding)的全部unigenes和差异表达基因数量也明显高于其他分子功能。值得注意的是,全部unigenes和差异表达基因GO功能分类的富集趋势并不完全相同。
根据千里光茎和叶中全部unigenes和差异表达基因的topGO前10位分类统计结果(表2),被注释为单生物细胞过程(single-organism cellular process,GO:0044763)、细胞代谢过程(cellular metabolic process,GO:0044237)以及大分子代谢过程(macromolecule metabolic process,GO:0043170)等10个功能的全部unigenes和差异基因数均较高,其中,被注释为生物过程(biological process,GO:0008150)的全部unigenes和差异表达基因的数量最多。
表1千里光茎和叶中全部unigenes和差异表达基因的GO功能分类
Table1GOfunctionclassificationofallunigenesanddifferentiallyexpressedgenesinstemandleafofSenecioscandensBuch.-Ham.exD.Don
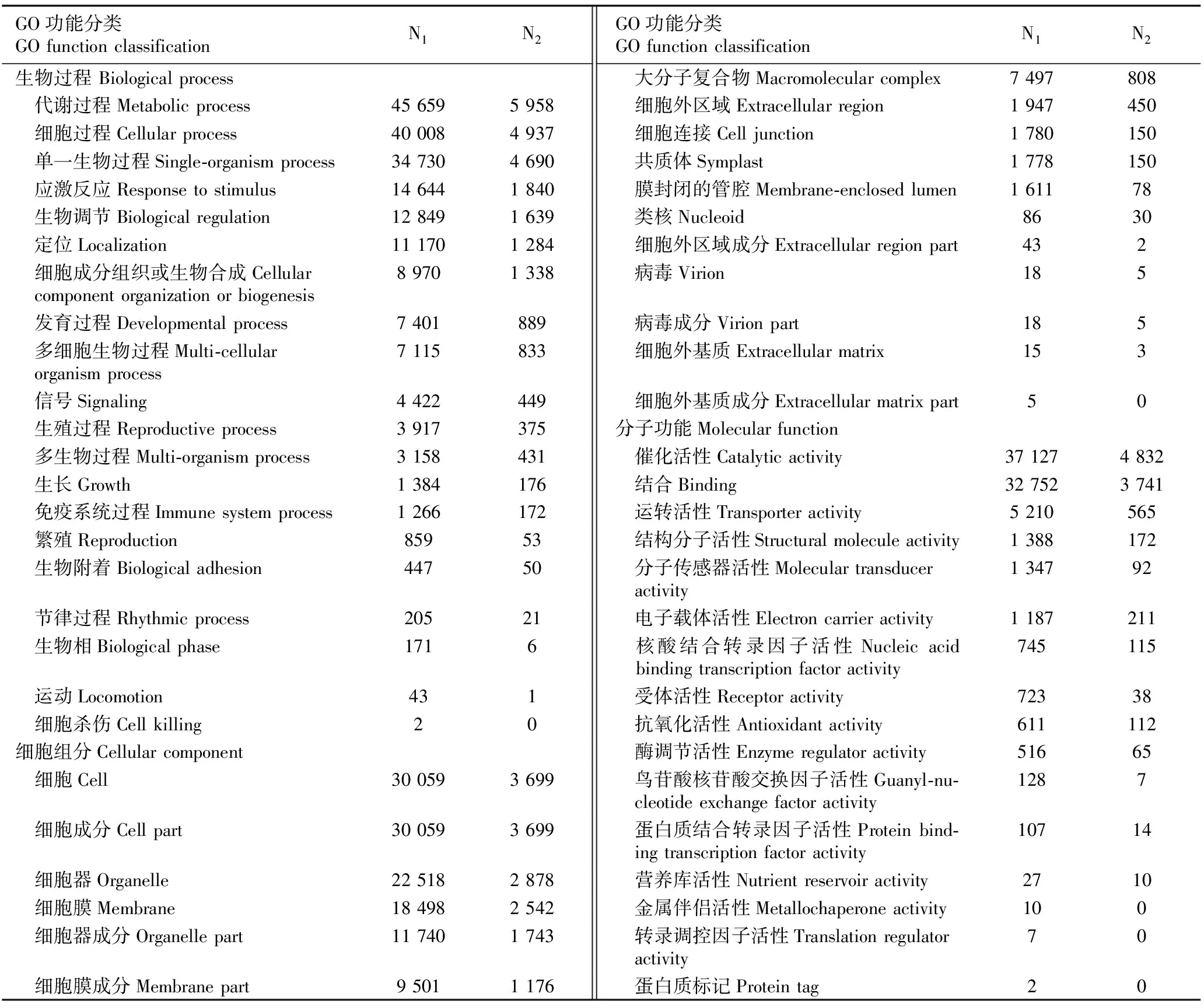
GO功能分类GO function classificationN1N2GO功能分类GO function classificationN1N2生物过程 Biological process 大分子复合物Macromolecular complex7 497808 代谢过程Metabolic process45 6595 958 细胞外区域Extracellular region1 947450 细胞过程Cellular process40 0084 937 细胞连接Cell junction1 780150 单一生物过程Single-organism process34 7304 690 共质体Symplast1 778150 应激反应Response to stimulus14 6441 840 膜封闭的管腔Membrane-enclosed lumen1 61178 生物调节Biological regulation12 8491 639 类核Nucleoid8630 定位Localization11 1701 284 细胞外区域成分Extracellular region part432 细胞成分组织或生物合成Cellular component organization or biogenesis8 9701 338 病毒Virion185 发育过程Developmental process7 401889 病毒成分Virion part185 多细胞生物过程Multi-cellular organism process7 115833 细胞外基质Extracellular matrix153 信号Signaling4 422449 细胞外基质成分Extracellular matrix part50 生殖过程Reproductive process3 917375分子功能Molecular function 多生物过程Multi-organism process3 158431 催化活性Catalytic activity37 1274 832 生长Growth1 384176 结合Binding32 7523 741 免疫系统过程Immune system process1 266172 运转活性Transporter activity5 210565 繁殖Reproduction85953 结构分子活性Structural molecule activity1 388172 生物附着Biological adhesion44750 分子传感器活性Molecular transducer activity1 34792 节律过程Rhythmic process20521 电子载体活性Electron carrier activity1 187211 生物相Biological phase1716 核酸结合转录因子活性Nucleic acid binding transcription factor activity745115 运动Locomotion43 1 受体活性Receptor activity72338 细胞杀伤Cell killing2 0 抗氧化活性Antioxidant activity611112细胞组分Cellular component 酶调节活性Enzyme regulator activity51665 细胞Cell30 0593 699 鸟苷酸核苷酸交换因子活性Guanyl-nu-cleotide exchange factor activity1287 细胞成分Cell part30 0593 699 蛋白质结合转录因子活性Protein bind-ing transcription factor activity10714 细胞器Organelle22 5182 878 营养库活性Nutrient reservoir activity2710 细胞膜Membrane18 4982 542 金属伴侣活性Metallochaperone activity100 细胞器成分Organelle part11 7401 743 转录调控因子活性Translation regulator activity70 细胞膜成分Membrane part9 5011 176 蛋白质标记Protein tag20
1)N1: 全部unigenes数量 Number of all unigenes; N2: 差异表达基因数量 Number of differentially expressed genes.
表2千里光茎和叶中全部unigenes和差异表达基因的topGO前10位分类统计结果
Table2StatisticalresultofthetoptenoftopGOofallunigenesanddifferentiallyexpressedgenesinstemandleafofSenecioscandensBuch.-Ham.exD.Don

编号IDGO功能分类GO function classificationN1N2GO:0044763单生物细胞过程Single-organism cellular process26 3393 481GO:0044237细胞代谢过程Cellular metabolic process32 2424 095GO:0044710单一生物代谢过程Single-organism metabolic process23 0143 388GO:0044699单一生物过程Single-organism process33 2594 465GO:0009987细胞过程Cellular process38 7614 885GO:0044238初级代谢过程Primary metabolic process31 8603 922GO:0043170大分子代谢过程Macromolecule metabolic process21 6762 453GO:0071704有机物代谢过程Organic substance metabolic process33 4994 308GO:0008152代谢过程Metabolic process39 7935 335GO:0008150生物过程Biological process49 4176 436
1)N1: 全部unigenes数量 Number of all unigenes; N2: 差异表达基因数量 Number of differentially expressed genes.
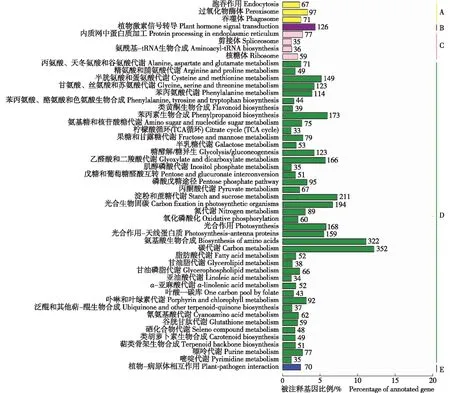
图中数据为差异表达基因数量 Data in the diagram indicate number of differentially expressed genes. A: 细胞过程 Cellular process; B: 环境信息处理 Environmental information processing; C: 遗传信息处理 Genetic information processing; D: 代谢 Metabolism; E: 生物体系 Organism system.
总体来看,这些差异表达基因与信号传导途径、次生代谢过程、细胞组分合成和酶催化活性等功能密切相关。
2.3.2 KEGG代谢通路分析 千里光茎和叶中差异表达基因的KEGG代谢通路分析结果(图2)表明:共有4 527个差异表达基因,涉及50个代谢通路,且大部分代谢通路属于代谢途径。其中,碳代谢途径的差异表达基因数量最多(352),占该通路差异表达基因总数的7.78%;其次为氨基酸生物合成途径,共有322个差异表达基因,占该通路差异表达基因总数的7.11%;淀粉和蔗糖代谢途径的差异表达基因数量也较多(211),占该通路差异表达基因总数的4.66%;而柠檬酸循环(TCA循环)途径的差异表达基因数量最少(33),占该通路差异表达基因总数的0.73%。
3 讨论和结论
由于药用植物各部位的药用成分含量和种类不同[13],因此,研究药用植物各器官基因的转录水平既利于阐明植物的器官分化基因,也利于深入分析药用成分积累的次生代谢途径及调控网络[14-16]。本研究从千里光茎和叶的cDNA文库中分别获得26 147 016和26 996 119个clean reads,Q30值分别为95.17%和94.74%,说明其转录组测序结果良好。千里光的茎中含有大量的木质素,木质素对维持较高的硬度及承托全株重量具有重要作用[17],而叶是植物合成及储存营养物质(如淀粉和蔗糖等)的主要场所[18]。本研究从千里光茎和叶中共筛选到10 991个差异表达基因,以茎中差异表达基因的表达量为准,叶中上调和下调的差异表达基因分别有5 542和5 449个;茎和叶中共有7 683个差异表达基因被GO功能注释成生物过程、细胞组成和分子功能3个大类53个亚类,且这些差异表达基因与信号传导途径、次生代谢过程、细胞组分合成和酶催化活性等密切相关。
KEGG代谢通路分析结果显示:千里光茎和叶中有4 527个差异表达基因,共涉及50个代谢通路,大部分差异表达基因参与碳代谢、氨基酸生物合成及淀粉和蔗糖代谢,说明这些差异表达基因与细胞组分合成和体内的生物合成等密切相关,其中,参与碳代谢和氨基酸合成的差异表达基因较多,据此推测千里光茎和叶的器官分化需要积累大量的多糖和蛋白质。千里光还含有丰富的次生代谢产物,包括生物碱类、酚酸类、黄酮类、挥发油类和萜类等[9],其中,类黄酮是千里光的主要抗菌成分之一[19],而供试植株SC-32具有强抗菌性[4],本研究发现参与类黄酮合成的差异表达基因有44个,说明这些差异表达基因主要参与千里光体内类黄酮的合成。据此认为,参与千里光茎和叶中类黄酮合成的差异表达基因可能在佐证类黄酮是千里光重要抗菌成分及千里光抗菌数量性状等方面具有重要作用。
综上所述,千里光茎和叶的差异表达基因具有器官特异性,其茎和叶的分化和形成与多糖和蛋白质积累密切相关。
