水蚀风蚀交错带退耕草坡地土壤酶活性和碳氮矿化特征
2019-03-25孔维波姚毓菲邵明安魏孝荣
孔维波, 石 芸, 姚毓菲, 邵明安, 魏孝荣
(1.中国科学院 教育部 水土保持与生态环境研究中心, 陕西 杨凌 712100; 2.陕西省定边县农技中心, 陕西 定边 718600;3.西北农林科技大学 黄土高原土壤侵蚀与旱地农业国家重点实验室, 陕西 杨凌 712100; 4.中国科学院大学, 北京 100049)
水蚀风蚀交错带是黄土高原生态环境最为脆弱的地区[1],该区由于长期开垦和不合理放牧,水土流失、草地退化和土壤沙化严重,生态环境受到极大破坏,是黄土高原退耕还林(草)措施实施的重点区域[2-3]。近年来在该区实施的退耕还草措施,使草地面积大幅增加,水土流失得到显著遏制,生态系统服务功能显著提升[4-5],对主要土壤过程产生重要影响。
水蚀风蚀交错带受水蚀和风蚀双重影响,相对于单一侵蚀营力下的生态系统环境更加脆弱,其对退耕措施的响应更为复杂[6]。在这一特殊区域开展退耕草地土壤碳氮过程研究,不但能够积累重要的基础资料,而且有利于理解复杂侵蚀环境下的植被演替与养分状况。该区退耕后草地演替顺序为:苜蓿—铁杆蒿—长芒草,两种草地群落在水蚀风蚀交错带分布普遍[7-8]。对不同演替阶段的草地群落土壤生物地球化学过程的认识,不但能够建立脆弱生态区退耕地演替阶段与养分循环过程之间的关系,而且可以为退耕草地主要土壤过程的科学评价和有效管理提供科学依据。
矿化作用是土壤碳氮循环的主要过程,不但对碳氮源汇有着重要的影响,而且决定着土壤养分的有效性和生态系统生产力水平[9],是土壤生物地球化学循环研究的重要内容之一。土壤酶活性对土壤生物过程具有敏感的指示作用,并且参与到土壤物质循环过程[10]。此外,由于测定方法较为成熟,并且相较于微生物指标的测定更易操作,土壤酶活性在土壤生物学研究中得到广泛应用[11]。目前研究者已经针对退耕条件下土壤酶活性与碳氮转化开展了大量的工作[12-13],但是很少有研究涉及到水蚀风蚀交错带和不同退耕阶段等因素,因此需要针对这些认识上的空白开展研究。
研究土壤性质空间分布特征,可以为退耕措施下土壤质量演变规律和机理的认识以及评价提供科学依据。目前在土壤理化性质空间分布方面已经进行了大量的工作,如王辽宏等[5]的研究表明,本氏针茅(Stipacapillata)坡地土壤—植物系统磷素在不同尺度上空间结构比较稳定,空间变异主要来源于结构性因素;胡伟等[14]发现,黄土高原退耕坡地土壤水分的相关距离随采样间距增大逐渐减小。但是很少有研究涉及到群落不同演替阶段对土壤酶活性及土壤碳氮矿化的空间异质性的影响,此外,土壤酶活性与碳氮矿化空间差异显著,具有高度空间变异性[15],而对这方面的认识,是正确理解土壤物质循环过程对不同管理措施或扰动响应机理的关键,也是在大的空间尺度上实现土壤碳氮循环及其效应预测的科学基础和重要手段。
坡地是土壤侵蚀发生的基本单元,是退耕措施实施的重点地貌类型[16]。本研究以黄土高原水蚀风蚀交错带不同演替阶段的退耕草坡地为对象,即苜蓿—铁杆蒿草地(Medicagosativa—Artemisiasacrorum)和长芒草草地(Stipabungeana),研究参与到土壤碳、氮和磷循环过程中的主要酶类以及碳氮矿化指标的变化特征,并应用地统计学方法分析这些指标在坡面的空间分布特征,以期为揭示不同演替阶段退耕草地的土壤生物学性质和碳氮循环过程及其响应机理提供科学依据。
1 材料与方法
1.1 研究区概况
本研究在位于神木县以西14 km的六道沟小流域进行。六道沟小流域位于黄土高原向毛乌苏沙漠、森林草原向典型干旱草原的过渡地带、北方农牧交错带和黄土高原水蚀风蚀交错带的核心区域。小流域年平均气温8.4℃,降水量437 mm,其中77%发生在6—9月份,属于中温带半干旱气候类型。植被类型为干旱草原,天然植被大部分遭到严重破坏,残存的天然草场也已退化、沙化,目前草地多为退耕地上自然演替发育的草地群落。该区地貌类型为片沙覆盖的黄土丘陵,坡地占整个流域的76.5%,地带性土壤为黑垆土,由于长期侵蚀的作用,已被黄绵土、新黄土、红土以及在沙地上发育起来的风沙土、坝地於土所取代。
1.2 样品采集与处理
本研究所选取的苜蓿—铁杆蒿草坡地坡长200 m,坡度约10°,长芒草草坡地坡长约150 m,坡度20°左右。于2007年8月在长芒草坡地沿坡顶向下每隔5 m设一个样带,共设30个样带;在苜蓿—铁杆蒿坡地沿坡顶向下每隔10 m设置一个样带,共设20个样带。每个样带内按“S”形布设5个样点,采集0—20 cm土壤样品并组成混合样品,带回实验室后,挑出植物残体和根系,过2.0 mm筛供土壤酶活性测定和碳氮矿化测定;另一部分土样过0.25 mm筛供主要养分含量(有机碳、全氮和全磷)测定。
1.3 土壤酶活性和碳氮矿化指标测定
土壤酶活性的测定方法和单位:土壤脲酶、蔗糖酶、碱性磷酸酶和淀粉酶活性分别采用Hoffmann与K.Teicher法、T.A.щербакова法、Ф.Х.Хазиев法和水杨酸比色法[17-18]。以上指标测定时均重复3次。
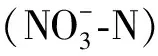
土壤养分指标测定了土壤有机质(重铬酸钾外加热法)、全氮(凯式蒸馏法)和全磷(HClO4-H2SO4消煮钼锑抗比色法)[17]。
1.4 地统计学分析
地统计学以区域化变量和随机函数为基础,以半方差函数为基本工具的统计方法。函数公式为:
(1)
式中:R(h)为变异函数值;N(h)为距离等于h时的点对总数;Z(xi)为区域化变量在位置xi处的实测数值;Z(xi+h)为在位置xi+h处的数值。目前用于拟合该函数模型有线性模型、球状模型、指数模型等,其中指数模型公式如下:
(2)
式中:R(h)在一定范围内随h增大而增大,到一定数值趋于稳定;C0为块金值,表示随机部分的空间异质性;C0+C为基台值,是半变异函数达到的极限值,值越大表示总的空间异质性越高。C0/(C0+C)为空间异质比,表示随机占总变异的比值,越大说明随机性变异越强,空间结构性越弱。一般的,比值<25%,20%~75%,>75%分别表示变量的空间相关性较强、中等、较弱[20]。
1.5 数据处理
采用一级动力学方程对有机碳累积矿化量进行拟合:
Cmin=Cp×(1-e-kt)
(3)
式中:Cmin为经过时间t(d)后土壤有机碳的累积矿化量(mg/kg);Cp为土壤潜在矿化量(mg/kg);k为有机碳矿化速率常数(d);t为培养时间(d)。
本研究统计分析均采用SPSS 19.0软件进行;半方差函数分析采用GS+9.0软件完成。
2 结果与分析
2.1 坡位和退耕草地类型对土壤养分的影响
本研究中主要土壤养分含量(有机碳、全氮和全磷)不受坡位(除苜蓿铁杆蒿坡地的全磷含量)影响(p>0.1)(表1,图1)。退耕草地类型对土壤养分含量的影响与养分种类有关,长芒草坡地有机碳含量(总体和下坡位)显著高于苜蓿—铁杆蒿坡地(高20.1%,36.5%,p<0.05,表1,图1A);群落类型对全氮和全磷含量影响不显著(p>0.1,表1)。此外,群落类型对有机碳和全氮含量的影响在不同坡位呈现出相似的趋势(p>0.05),但是对全磷含量的影响则与坡位有关(p<0.05)。苜蓿—铁杆蒿坡地全磷含量在上、中坡位分别高于长芒草坡地(高14.4%,18.0%),而在下坡位则显著低于长芒草坡地(低20.2%)。
2.2 坡位和退耕草地类型对土壤酶活性的影响
总的来说,坡位显著影响土壤酶活性(表1),随坡位的降低,脲酶和淀粉酶活性分别从25.86,31.05 μg/(g·h)增大到38.54,37.10 μg/(g·h)(p<0.05,图2A,2D),而蔗糖酶和碱性磷酸酶活性则分别从688.23,83.33 μg/(g·h)降低到494.00,67.14 μg/(g·h)(p=0.06)(图2B,2C)。群落类型对土壤酶活性的影响与酶类型有关,苜蓿—铁杆蒿坡地土壤蔗糖酶和碱性磷酸酶活性分别比长芒草坡地高187.57,22.46 μg/(g·h)(p<0.01,表1)。总体来说,土壤脲酶和淀粉酶活性不受群落类型的影响(p>0.1,表1),如苜蓿—铁杆蒿和长芒草坡地土壤脲酶活性分别为31.95 μg/(g·h)[28.92~34.75 μg/(g·h)],35.42 μg/(g·h)[22.81~44.91 μg/(g·h)],淀粉酶活性分别为31.99 μg/(g·h)[29.95~34.13 μg/(g·h)],34.57 μg/(g·h)[30.20~40.06 μg/(g·h)]。此外,群落类型对碱性磷酸酶和淀粉酶活性的影响在不同坡位呈现出相似的趋势(p>0.1),但是对脲酶和蔗糖酶活性的影响则与坡位有关(p<0.05)。如苜蓿—铁杆蒿坡地脲酶活性在上坡位显著高于长芒草坡地(高26.8%),而在下坡位则显著低于长芒草坡地(低28.4%);蔗糖酶活性在上坡位和中坡位显著高于长芒草坡地(p<0.05,分别高89.1%,46.8%),在下坡位则低于长芒草坡地(p<0.05,低19.05%)(图2A,2B)。
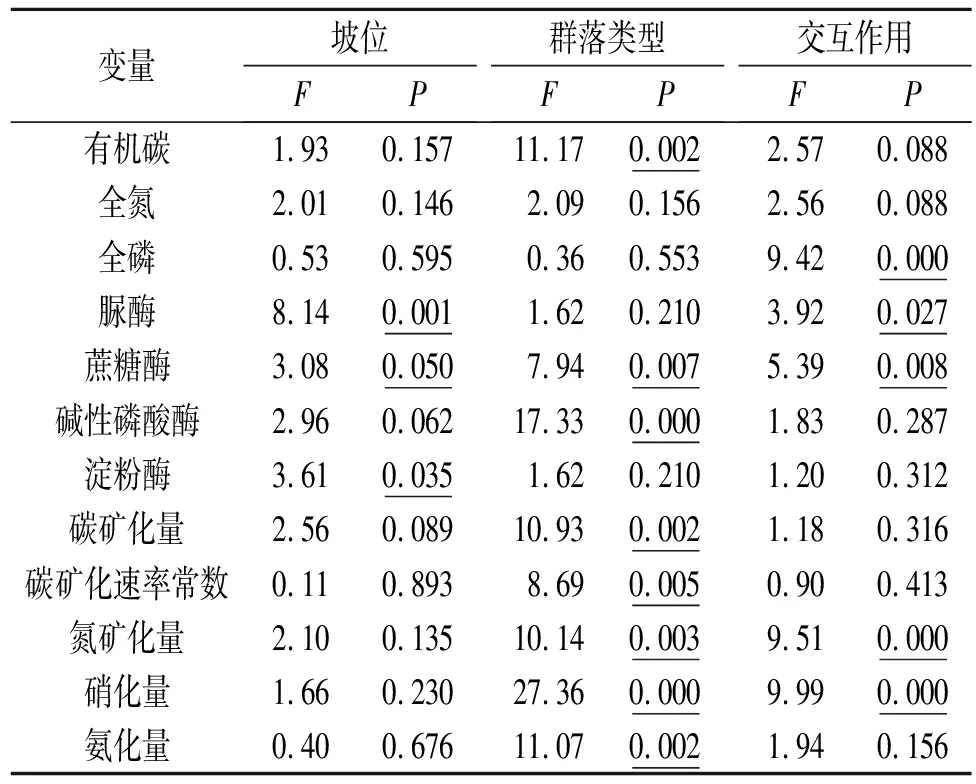
表1 坡位和草地类型对土壤养分含量、酶活性和碳氮矿化指标影响的方差分析结果
注:表中下划线数字表示统计显著。

注:不同大、小写字母分别表示土壤养分含量在长芒草和苜蓿铁杆蒿坡地不同坡位差异显著,*表示该指标在不同草地类型间差异显著(p<0.05);误差线为均值的标准误。
图1长芒草坡地和苜蓿-铁杆蒿坡地不同坡位的土壤养分含量
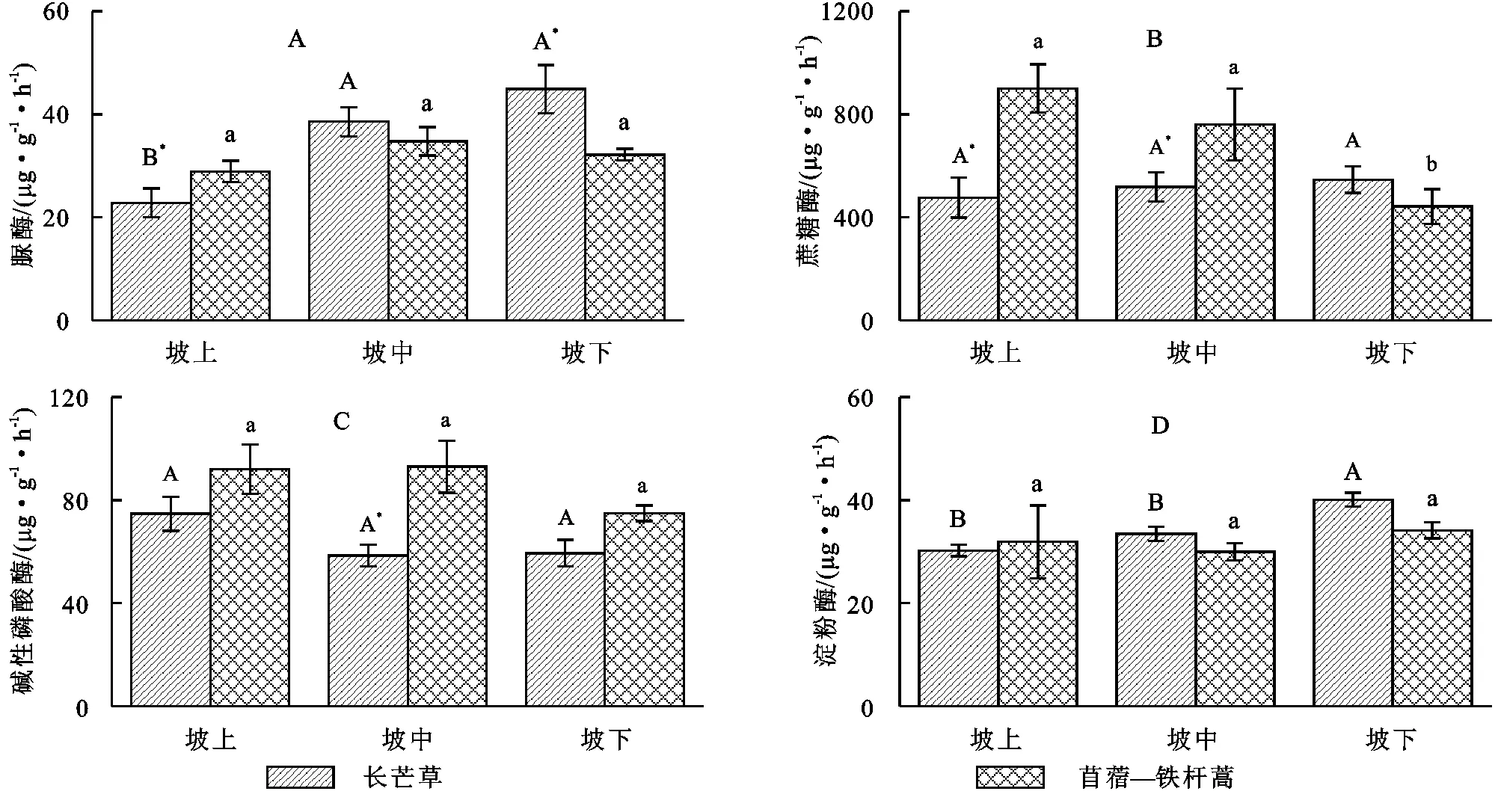
注:不同大、小写字母分别表示土壤酶活性在长芒草和苜蓿铁杆蒿坡地不同坡位差异显著,*表示该指标在不同草地类型间差异显著(p<0.05);误差线为均值的标准误。
图2长芒草坡地和苜蓿-铁杆蒿坡地不同坡位的土壤酶活性
2.3 坡位和退耕草地类型对土壤碳氮矿化特征的影响
土壤有机碳矿化量与矿化速率常数不受坡位影响(p>0.09)(表1),长芒草和苜蓿—铁杆蒿坡地有机碳矿化量分别介于116.98~135.76 mg/kg,108.23~118.11 mg/kg之间,每日矿化速率常数分别介于0.22~0.26,0.16~0.18之间。长芒草坡地有机碳矿化量和矿化速率常数在所有坡位均高于苜蓿—铁杆蒿坡地(下坡位显著,p<0.05),如有机碳矿化量在上、中、下坡位分别比苜蓿—铁杆蒿坡地高10.4%,8.1%,22.8%,矿化速率常数分别高19.5%,58.2%,23.8%(图3A,3B)。
研究区土壤氮素矿化由硝化作用主导,而氨化量呈现出负值,表明土壤微生物对铵态氮有较强的固定作用[21](图3D,3E)。苜蓿—铁杆蒿坡地氮素矿化量和硝化量显著高于长芒草坡地(p<0.01),但是这种影响与坡位有关(p<0.01)(表1),如苜蓿—铁杆蒿坡地氮矿化量在上坡位和中坡位分别比长芒草坡地高368.4%,150.5%,但是在下坡位则比长芒草坡地低50.2%;硝化量在上坡位和中坡位分别比长芒草坡地高204.9%,55.0%,在下坡位则低2.2%(图3D)。苜蓿—铁杆蒿坡地土壤氨化量显著低于长芒草坡地(p<0.01),而且这种影响与坡位无关(p>0.1),如氨化量在上、中和下坡位分别比长芒草坡地低92.8%,6.6%,75.2%(图3E)。这些结果表明,土壤碳氮矿化主要指标在不同坡位对退耕草地类型呈现出相似的响应特征,因此退耕后土壤碳氮矿化主要受退耕草地类型的影响。
2.4 退耕草地土壤酶活性和碳氮矿化指标的空间变异特征
退耕草坡地土壤酶活性空间变异的最佳拟合模型与草地类型和酶种类有关(表2)。脲酶、蔗糖酶、碱性磷酸酶和淀粉酶最佳拟合模型在长芒草坡地分别为指数、球状、球状和高斯模型,在苜蓿—铁杆蒿坡地分别为球状、高斯、高斯和指数模型(表2),表明土壤酶活性空间结构特征受草地群落类型的影响。与苜蓿—铁杆蒿坡地相比,长芒草坡地脲酶和淀粉酶活性空间变异的基台值较高(分别高110%,283%),空间变异较大;蔗糖酶和碱性磷酸酶活性的基台值较低(分别低38.2%,54.9%),空间变异较小。此外,长芒草坡地4种土壤酶活性具有较强的空间自相关性(C0/C0+C<25%),变异主要由结构因素引起。苜蓿—铁杆蒿坡地蔗糖酶和碱性磷酸酶活性空间自相关性中等(C0/C0+C介于25%~75%),变异由随机因素和结构因素引起;脲酶和淀粉酶活性具有较强的空间自相关性(C0/C0+C<25%),变异由结构因素导致。长芒草坡地脲酶和淀粉酶活性的变程(分别为633,208 m)大于苜蓿铁杆蒿坡地(分别为10,21 m),具有较大的自相关范围表明随着苜蓿—铁杆蒿群落向长芒草群落的演替,脲酶和淀粉酶活性的空间自相关性增强;蔗糖酶和碱性磷酸酶呈相反的变化趋势,空间自相关性减弱,但仍然具有强的空间自相关性。
有机碳累积矿化量和矿化速率常数、氮素矿化量和氨化量空间变异最佳拟合模型在长芒草坡地均为球状模型,在苜蓿—铁杆蒿坡地则分别为高斯、线性、指数和球状模型,表明这4类指标的空间结构特征也受草地群落类型的影响(表2)。长芒草坡地有机碳矿化量、氮素矿化量和氨化量具有强烈的空间自相关性(C0/C0+C<25%),变异由结构因素引起;矿化速率常数具有中等空间自相关性(C0/C0+C介于25%~75%),变异由随机因素和结构因素共同引起。苜蓿—铁杆蒿坡地有机碳和氮的矿化量具有中等空间自相关性(C0/C0+C介于25%~75%),变异由随机因素和结构因素共同引起,有机碳矿化速率常数具有较弱的空间自相关性(C0/C0+C>75%),氨化量则具有强烈的空间自相关性(C0/C0+C<25%)。此外,长芒草坡地4种指标的变程均小于苜蓿—铁杆蒿坡地。上述结果表明,随着苜蓿—铁杆蒿群落向长芒草群落的演替,碳氮矿化指标的空间自相关性逐渐增强。
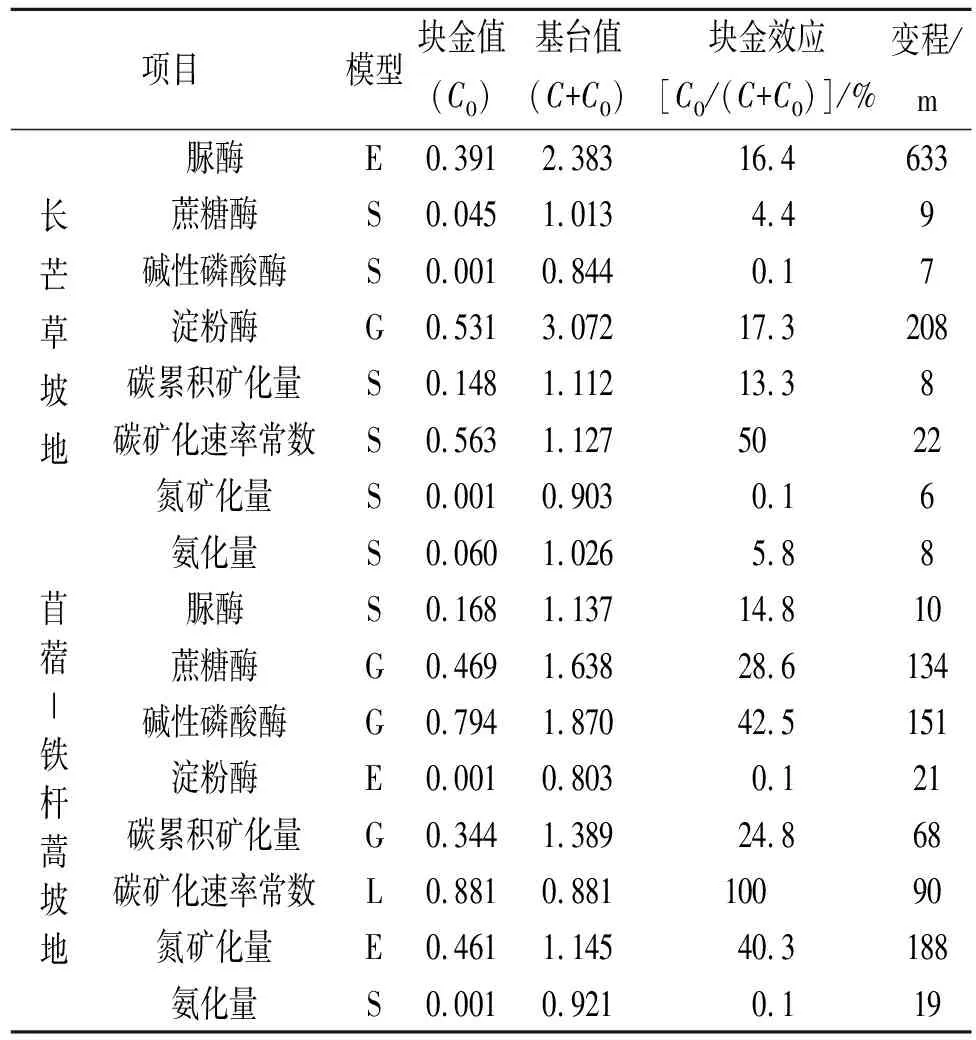
表2 不同草地土壤酶活性和碳氮矿化指标的半方差理论及模型参数
注:G代表高斯模型,S代表球状模型,E代表指数模型。
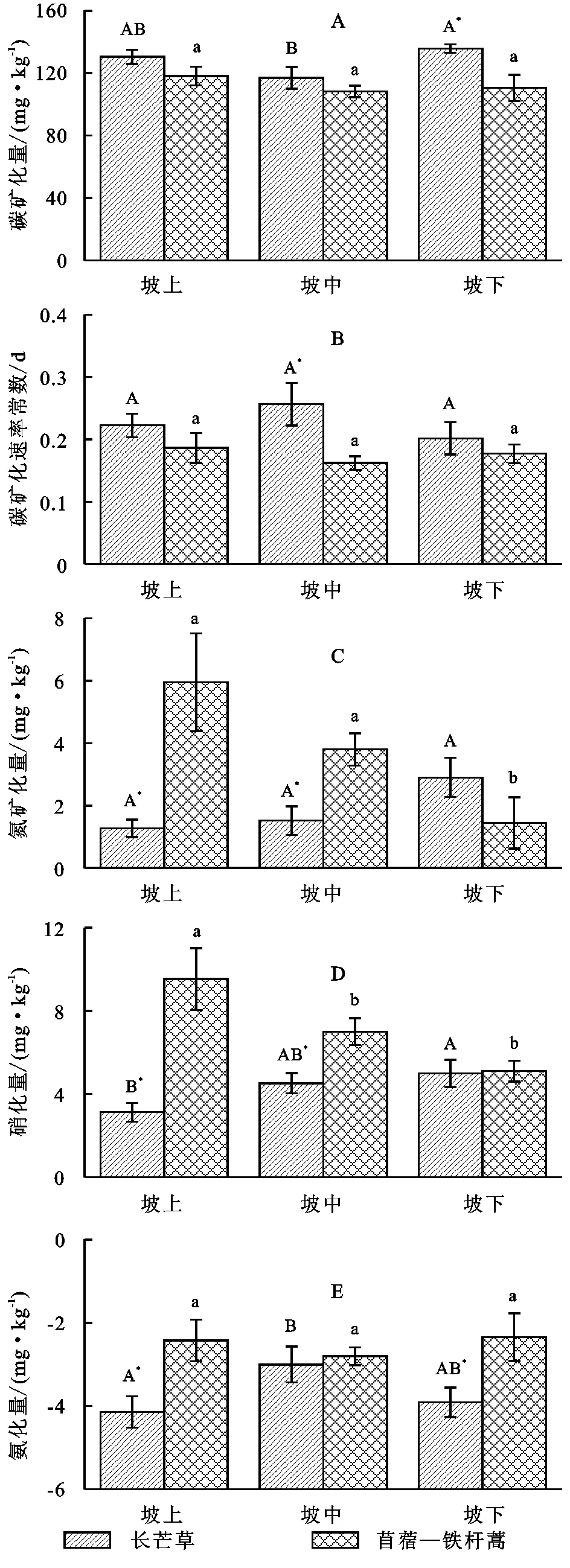
注:不同大、小写字母分别表示土壤碳氮矿化指标在长芒草和苜蓿铁杆蒿坡地不同坡位差异显著,*表示该指标在不同草地类型间差异显著(p<0.05);误差线为均值的标准误。
图3长芒草坡地和苜蓿-铁杆蒿坡地不同坡位的土壤碳氮矿化指标
3 讨论与结论
3.1 土壤养分分布
草地群落演替显著改变了物种组成和群落生产力,并对土壤养分的有效性、植物吸收、植物养分向土壤的归还过程等产生影响,从而改变土壤养分的空间分布和动态特征[22]。此外,这些过程对群落演替的响应还与地形条件有关[23]。
本研究结果表明,长芒草坡地有机碳和全氮含量均高于苜蓿—铁杆蒿坡地,这是因为不同植被类型存在土壤养分含量的差异[24],且随着草地演替的进行,草地质量得以提高[25],如张婷等[7]发现的黄土丘陵区长芒草群落有机质和全氮含量均显著高于铁杆蒿群落的结果相一致。长芒草坡地全磷含量在上坡位和中坡位低于苜蓿—铁杆蒿坡地(图1),可能与植被演替阶段有关,长芒草为退耕后植被演替的顶级群落,演替时间长于苜蓿—铁杆蒿群落,植被从土壤中吸收的磷总量较高,因此造成土壤中全磷含量的降低。但是长芒草坡地土壤全磷含量在下坡位显著高于苜蓿—铁杆蒿坡地,可能与坡面水土过程造成的磷素迁移有关[2]。并且长芒草坡地坡度大于苜蓿—铁杆蒿坡地,其坡面水土过程造成的土壤颗粒迁移和在下坡位的沉积作用强于苜蓿—铁杆蒿坡地,从而造成了两个类型坡地下坡位全磷含量的差异。以上表明,在评估恢复坡地生态系统土壤养分动态和有效性方面,需要考虑群落类型和坡位的影响。
3.2 土壤酶活性
坡位显著影响土壤酶活性,随坡位的降低,脲酶和淀粉酶活性升高,蔗糖酶和碱性磷酸酶活性降低,而成毅等[26]发现的土壤酶活性不随坡位的变化而变化。这种差异可能与土壤酶活性受到多种因素的影响有关(土壤pH值、生长季、气候条件、生物学特征和微地形的变化等)[27-28]。草地群落的演替可以改变土壤性质,包括有机质形态、土壤pH值和养分有效性等[29];同时不同植被根系分布及其分泌物和营养吸收方式的不同也影响着微生物生物量[30]。而土壤酶主要来源于植物和微生物(细菌和真菌)活动,其功能与活性和微生物联系紧密[29],因此,退耕草地类型(或者演替过程)对土壤酶活性产生显著影响,而且这种影响与土壤酶类型有关[31]。本研究中长芒草坡地脲酶、淀粉酶活性高于苜蓿—铁杆蒿坡地,这与安韶山等[32]的研究一致;蔗糖酶活性小于苜蓿—铁杆蒿坡地[33],与安韶山等[32]的研究相反,可能是由于土壤酶活性不仅与植被盖度、多度、高度有关,同时还取决于当地的气候和土壤理化性质[34-35]。
3.3 土壤碳氮矿化特征
土壤有机碳矿化受多种因素的影响,如土壤有机碳含量[36]、有机质组分和存在状态[37]、土壤理化性质[38]、微生物群落组成[39]等。本研究中坡位对长芒草和苜蓿—铁杆蒿坡地有机碳矿化没有显著影响(p>0.09),可能与不同坡位土壤有机碳含量差异不显著有关,有机碳矿化与土壤有机碳含量显著正相关性也证明了此结果(R2=0.423,n=50,p<0.002)。虽然有研究表明,氮素矿化与全氮和有机碳含量呈显著正相关系,且本研究中长芒草坡地有机碳和全氮含量高于苜蓿铁杆蒿坡地,但是氮素矿化却呈现苜蓿—铁杆蒿坡地显著高于长芒草坡地,可能是由于土壤氮素矿化受植物氮素含量(主要是根系氮素含量)驱动[40],豆科植物(如苜蓿)根系生物固氮,能显著促进土壤氮素矿化[41]。长芒草群落氮矿化量下坡位最高[4],苜蓿—铁杆蒿群落矿化量下坡位最低[2],可能由于长芒草群落土壤酶活性在下坡位较高,促进土壤氮素矿化[42],而苜蓿—铁杆蒿群落土壤酶活性在坡位之间没有显著差异,并且苜蓿—铁杆蒿坡地不同坡位群落物种不同(上坡位以苜蓿为主,中坡位二者共同主导,下坡位铁杆蒿为主)[2],从而对氮素矿化产生影响[43]。因此,不同类型坡地土壤氮素矿化呈现出如图3所示的随坡位变化趋势。土壤硝化量与矿化量极显著正相关(R2=0.87,n=50,p<0.01),因此,苜蓿—铁杆蒿坡地硝化量也以下坡位最低,而长芒草坡地硝化量也以下坡位最高(图3)。此外,氮的微生物固定导致的研究中土壤氨化量呈负值[24],且不同坡位差异不显著,表明微生物对铵的固定不受坡位影响,研究区氮素矿化以硝化作用主导。
3.4 土壤酶活性和碳氮矿化指标的空间变异特征
土壤酶活性的空间结构特征受群落类型的影响,长芒草坡地4种酶活性具有较强的空间自相关性,其变异主要由结构性因素引起(表2),这一结果与朱新萍等[44]对高产棉田土壤脲酶和碱性磷酸酶活性空间自相关性的研究结果,以及陈立明等[15]发现的不同退化程度云冷杉林土壤酶活性的空间变异主要由结构因素引起的研究结果一致。本研究中苜蓿—铁杆蒿坡地脲酶和淀粉酶活性具有较强的空间自相关性,蔗糖酶和碱性磷酸酶活性具有较弱的空间自相关性,表明不同类型土壤酶活性的空间变异特征差异较大,这与土壤酶的专一特性有关。不同坡地土壤酶活性空间变异特征的差异,还与土壤理化性质空间变异特征有关。张燕江等[2]对研究区退耕坡地苜蓿—铁杆蒿群落养分分布特征的研究发现,随着人工草地向天然草地的演替,群落养分含量的空间结构性逐渐增强,而土壤酶活性与这些性质密切相关,因此随着苜蓿—铁杆蒿坡地向长芒草坡地的演替,土壤酶活性的空间结构逐渐增强。
本研究中苜蓿—铁杆蒿坡地土壤碳氮矿化指标具有中等空间自相关性,变异主要由结构因子和随机因子共同造成,长芒草坡地碳氮矿化指标具有强烈的空间自相关性,变异主要由结构性因素引起,表明土壤碳氮矿化指标的空间结构性随着群落演替的进行逐渐增强,这与其他研究结果相一致,如朱剑兴等[45]发现,土壤碳氮矿化指标随草地演替(过渡草地向典型草地)呈现出很好的空间格局。此外,坡地土壤对地表水土过程和植被生长与分布响应敏感,这些过程均使土壤资源重新分配并对养分转化指标的空间格局产生影响。Wei等[46]的研究结果表明,研究区天然草坡植被覆盖相对均匀,坡面土壤资源的重新分配作用较弱,而退耕坡地植物覆盖不均,土壤资源沿坡面的重新分配作用和空间异质性较强,而土壤碳氮矿化受土壤有机质含量与组成、环境因子和群落组成的影响,因此碳氮矿化指标的空间结构呈现出随苜蓿铁杆蒿坡地向长芒草坡地的演替增强的趋势(表2)。
