硫素形态对苗期菘蓝生长生理特性及次生代谢的影响
2019-03-23缪雨静关佳莉曹艺雯沙俊涛唐晓清耿丽王磊
缪雨静,关佳莉,曹艺雯,沙俊涛,唐晓清*,耿丽,王磊
(1.南京农业大学中药材研究所,江苏 南京 210095; 2.句容市茅山仙草中药材专业合作社,江苏 句容 212404)
硫作为植物所需中量元素之一,约占植株干重的0.1%,虽不是植物细胞的结构性组成元素,但它却发挥极其重要的作用[1]。硫素是一些氨基酶的组成成分,也是蛋白质合成的必须元素,有助于酶和维生素的形成。同时,硫素还参与植物光合作用,为叶绿素合成所必需[2]。因此,硫素在植物生长、产量形成以及初次生物质积累过程中发挥着重要作用。
植物对硫元素的吸收,除从土壤中获得硫酸盐外,还能直接吸收大气中的SO2,此外,植物还可用根系和叶片吸收S2-、HSO3-和SO32-等形态的无机硫素以及有机含硫化合物作为营养,以保证其正常的生长和发育[3]。硫素被吸收后,会在植物体内经过一系列的还原同化,最终才能被植物体所利用。植物中的硫酸盐同化主要包括3个阶段:活化阶段、还原阶段和半胱氨酸(cysteine, Cys)合成阶段[4-5]。SO42-化学性质稳定,首先会通过ATP-硫酸化酶(ATP sulfurylase, ATPS)和APS磺基转移酶(APS reductase, APR)氧化形成SO32-,然后经硫化物还原酶形成硫化物(S2-),最后在半胱氨酸合成酶(cysteine synthase, OAS-TL)的催化下形成半胱氨酸,其为合成植物体内蛋白质和氨基酸酶的来源[6]。因此,不同形态的硫素可能参与到同化过程的特定阶段,对植物硫素吸收形成一个反馈作用,从而影响植物对硫元素的吸收和利用。此外,由于硫酸盐与硝酸盐同化是协调进行的,所以不同的硫素形态同样会影响植物对氮素的吸收同化及利用。对于需硫量较大的十字花科植物而言[7-8],硫素在次生代谢过程中也起着至关重要的作用,如十字花科植物中的芥子油苷合成过程中伴随着硫的同化,硫吸收同化产物可直接应用于芥子油苷的合成途径中[9]。在菘蓝(Isatisindigotica)中,氮硫素配施对于叶片靛蓝和靛玉红的含量大致呈现为高低硫抑制,适宜硫浓度促进的趋势[10]。
菘蓝是十字花科菘蓝属两年生草本植物,根和叶均可入药。叶入药为大青叶,根入药为板蓝根,具有清热解毒、凉血等功效。两者均为我国常用的清热解毒之要药[11]。目前对于菘蓝的营养研究主要集中在氮素营养方面,而硫元素对菘蓝生长的影响尚无系统研究。因此,本研究通过分析不同硫素形态对菘蓝生长、光合生理、氮硫代谢及次生物质积累的影响,寻找适宜提高菘蓝药材品质的硫素形态,旨在为合理使用硫素肥料及提高大青叶品质提供理论基础和应用参考。
1 材料与方法
1.1 材料培养
供试材料菘蓝种子为来自内蒙古的栽培居群,经南京农业大学王康才教授鉴定为十字花科菘蓝属植物菘蓝的角果(生产中称种子)。试验于2017年8-12月在南京农业大学园艺学院阳光温室内进行,以蛭石和珍珠岩(2∶1)作为栽培基质进行育苗,于4叶期进行间苗,每盆留10株长势一致的菘蓝幼苗,进行后期Hoagland营养液浇灌处理。
1.2 试验设计
试验设置6个处理,分别为Na2SO4,Na2SO3,NaHSO3,Na2S,Na2S2O35种不同形态的硫素,并以无硫作为对照处理(CK)。各处理施以改良的Hoagland营养液,微量元素和铁元素参考Zheng等[12]的研究。育苗期施1/4 Hoagland(无硫)营养液,处理期施添加不同形态硫素的Hoagland营养液,硫素浓度统一为2.5 mmol·L-1,每5 d浇灌一次,处理45 d后,进行采样,鲜样叶片冻于-80 ℃冰箱内保存,干样叶片烘干至恒重,打粉,待用。
1.3 测定项目与方法
1.3.1生物量测定 随机选取10株菘蓝,洗净拭干后于105 ℃杀青15 min后于60 ℃烘干至恒重,测量菘蓝地上部位(指茎基部至最长叶片之间的距离称株高)长度,并称其鲜干重,叶片烘干粉碎后过四号筛(250 μm),用于活性成分含量测定。
1.3.2光合参数测定 测定前提前将菘蓝放置阳光下30 min充分适应,于晴朗少云上午9-11时,选取菘蓝植株由内至外的第二轮功能叶,用 LI-6400 型光合仪(美国LI-COR公司)于1000 μmol·m-2·s-1光强下,测定菘蓝叶片净光合速率(net photosynthetic rate,Pn)、气孔导度(stomatal conductance,Gs)、胞间二氧化碳浓度(intercellular CO2concentration,Ci)和蒸腾速率(transpiration rate,Tr),以及计算水分利用率(water use efficiency, WUE)每个处理5次重复。
1.3.3光合色素含量测定 参照王学奎等[13]的方法测定菘蓝叶片的光合色素含量。称取0.05 g叶片剪成碎片,置于10 mL 95%乙醇内,避光浸提至叶片为无色。用紫外分光光度法于665,649和470 nm下测定吸光值。根据公式计算叶绿素a,叶绿素b,类胡萝卜素和叶绿素总量,各处理3次重复。计算公式如下:Ca=13.95×A665-6.88×A649;Cb=24.96×A649-7.32×A665;Cx·c=(1000×A470-2.05×Ca-114.8×Cb)/245。
1.3.4氮代谢酶和硫代谢酶活性测定 硝酸还原酶(nitrate reductase, NR)、谷氨酰胺合成酶(glutamine synthetase, GS)、谷氨酸脱氢酶(glutamate dehydrogenase, GDH)、谷氨酸合成酶(glutamate synthetase, GOGAT)、ATP-硫酸化酶、APS磺基转移酶和半胱氨酸合成酶活性采用南京建成生物研究所试剂盒测定,各处理3次重复。
1.3.5可溶性糖、可溶性蛋白和游离氨基酸含量测定 采用苯酚硫酸法测定可溶性糖含量,考马斯亮蓝G-250法测定可溶性蛋白含量,酸性茚三酮法测定游离氨基酸含量[13]。
1.3.6靛蓝、靛玉红含量测定 参照2015版《中国药典》方法提取、测定靛蓝和靛玉红含量,略作修改[11]。超高效液相色谱(ultra performance liquid chromatography, UPLC)条件:分析柱为Aglient ZORBA×Eclidise Plus C18 (2.1 mm×50 mm,1.8 μm);流动相为甲醇∶水(72∶28,v/v); 流速 0.30 mL·min-1;柱温30 ℃;检测波长 289 nm;进样体积 2 μL。分别以靛蓝、靛玉红的色谱峰面积(Y)与其对应的含量(X,μg·mL-1)作标准曲线,计算回归方程。靛蓝的回归方程为:y=16.022x+21.742,R2=0.9994(n=3),线性范围:0~2 μg·mL-1;靛玉红的回归方程为:y=157.32x-9.3294,R2=0.9998(n=3),线性范围:0~10 μg·mL-1。
1.4 数据统计与分析
采用Microsoft Excel 2010 和SPSS 19.0软件进行数据整理和统计分析,结果以“平均值±标准误”表示。
2 结果与分析
2.1 不同硫素形态对苗期菘蓝生长及生物量的影响
表1结果显示,在不同硫素形态下菘蓝生长存在差异。其中NaHSO3和Na2SO4处理下菘蓝株高显著高于其他硫素形态处理,分别为25.46和24.80 cm。Na2S处理下菘蓝苗株高最低,分别低于其他处理8.40%(CK),21.69%(Na2SO4),7.30%(Na2SO3),23.72%(NaHSO3),22.81% (Na2S2O3)。菘蓝叶片鲜重以Na2SO4处理最高,其余处理依次为NaHSO3>CK>Na2SO3>Na2S2O3>Na2S。干重也以Na2SO4处理最高,分别高于其他处理66.67% (CK),87.50%(Na2SO3),18.81%(NaHSO3),179.07%(Na2S),64.38%(Na2S2O3)。综合分析,以Na2SO4处理最有利于菘蓝生长及生物量的积累。
2.2 不同硫素形态对苗期菘蓝光合参数的影响
由表2可以看出,净光合速率变化趋势与水分利用效率基本一致,以NaHSO3处理下最高,且与其他处理呈显著差异(P<0.05)。Na2SO4处理次之,而Na2S2O3处理处于最低水平。气孔导度以NaHSO3处理最高,Na2S2O3次之,其余处理间无显著差异(P>0.05)。蒸腾速率与气孔导度变化存在一定的相似性,依次为NaHSO3>Na2S2O3>Na2SO3>Na2SO4>CK>Na2S。胞间CO2浓度变化与净光合速率变化趋势相反, NaHSO3处理下胞间CO2浓度最低,而净光合速率最小的Na2S2O3处理的胞间CO2浓度最高。综合分析NaHSO3处理最有利于菘蓝叶片光合作用,Na2SO4处理次之。

表1 不同硫素形态对苗期菘蓝生长及生物量的影响 Table 1 Effect of different sulfur forms on growth, leaf fresh and dry weight of I. indigotica (n=10)
注:表中数值后小写字母表示处理间差异达0.05显著水平,下同。
Note:Values in the table followed by different letters are significant at 0.05 level, the same below.

表2 不同硫素形态对苗期菘蓝光合参数的影响 Table 2 Effect of different sulfur forms on photosynthetic parameters of I. indigotica (n=5)
2.3 不同硫素形态对苗期菘蓝光合色素的影响
从表3可知,不同形态硫素处理对菘蓝苗期叶绿素总量和叶绿素a含量并无显著影响(P>0.05)。叶绿素b含量以Na2S处理最高,分别高于其他处理58.97%(CK),34.78%(Na2SO4),51.22%(Na2SO3),34.78%(NaHSO3),58.97%(Na2S2O3)。而类胡萝卜素以Na2S处理最低,与其余各处理差异显著(P<0.05),而其余处理间并无显著差异(P>0.05)。
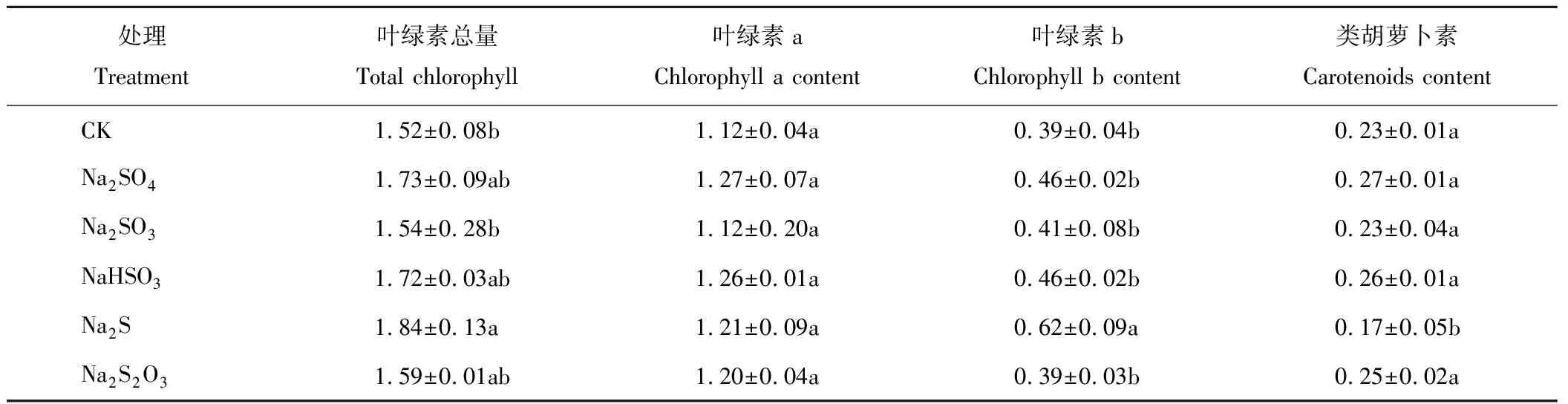
表3 不同硫素形态对苗期菘蓝光合色素含量的影响 Table 3 Effect of different sulfur forms on photosynthetic pigment contents of I. indigotica (n=3) (mg·g-1)
2.4 不同硫素形态对苗期菘蓝氮代谢酶活性的影响
在不同硫素形态条件下, Na2SO3处理叶片中硝酸还原酶活性最高,与其他处理差异显著(P<0.05)(表4)。CK和Na2SO4处理后硝酸还原酶活性相比于Na2SO3分别下降了20.44%和22.22%,两者无显著差异(P>0.05)。谷氨酰胺合成酶活性于对照处理(CK)下最高,分别高于其他处理34.39%(Na2SO4),41.56%(Na2SO3),61.50%(NaHSO3),20.24%(Na2S),23.34%(Na2S2O3)。谷氨酸脱氢酶以Na2SO4处理最低,且相较于其他处理,分别下降了1.48(CK),2.24(Na2SO3),0.60(NaHSO3),2.66(Na2S),3.79(Na2S2O3)倍。谷氨酸合成酶以Na2S处理最高,CK、Na2SO4、Na2S2O3次之,且3者间无显著差异(P>0.05)。SO32-形态的硫素(Na2SO3和NaHSO3)处理后谷氨酸合成酶活性最低。综上发现,硫素形态对氮代谢酶活性影响没有明显的规律。

表4 不同硫素形态对苗期菘蓝氮代谢酶活性的影响 Table 4 Effect of different sulfur forms on nitrogen metabolism enzymes of I. indigotica (n=3)
2.5 不同硫素形态对苗期菘蓝硫代谢酶活性的影响
表5可以看出,不同形态的硫素对苗期菘蓝叶片中硫代谢酶活性均以Na2SO4处理为最高(ATPS除外),且显著高于其他处理(P<0.05)。其中ATPS活性以CK处理最高,Na2SO4次之,但两者间无显著差异(P>0.05)。Na2SO4处理分别比其他处理上升了85.79%(Na2SO3),193.21%(NaHSO3),47.25%(Na2S),156.40%(Na2S2O3)。APR活性分别是其他处理的2.70(CK),2.23(Na2SO3),3.36(NaHSO3),3.12(Na2S),5.36(Na2S2O3)倍。OAS-TL活性在NaHSO3、CK和Na2S处理间差异不显著,Na2SO3和Na2S2O3处理间差异不显著。由此可见,以Na2SO4处理下菘蓝叶片中硫代谢酶活性最强。

表5 不同硫素形态对苗期菘蓝硫代谢酶活性的影响 Table 5 Effect of different sulfur forms on sulfur metabolism enzymes of I. indigotica (n=3)
2.6 不同硫素形态对苗期菘蓝中营养物质含量的影响
图1结果显示,不同形态硫素对苗期菘蓝叶片中可溶性糖、游离氨基酸和可溶性蛋白含量的影响趋势不同。其中可溶性糖含量以NaHSO3处理最高,与其他处理差异显著(P<0.05), Na2SO3和Na2SO4处理次之, CK、Na2S和Na2S2O3处理处于较低水平。施硫肥处理组游离氨基酸含量均高于无硫处理组(CK)。其中以Na2SO4处理下游离氨基酸含量处于最高水平。与游离氨基酸类似,可溶性蛋白含量于无硫处理下(CK)最低,相比于其他处理分别下降了56.07%(Na2SO4),55.52%(Na2SO3),81.94%(NaHSO3),78.89%(Na2S)和79.72%(Na2S2O3)。其中NaHSO3处理的可溶性蛋白含量最高。
2.7 不同硫素形态对苗期菘蓝中靛蓝和靛玉红含量的影响
不同形态硫素条件下菘蓝叶片中活性成分含量变化与生理变化存在一定的差异(图2)。靛蓝和靛玉红含量均以Na2S处理最高。其中靛蓝含量以CK、Na2SO4、Na2SO3和Na2S2O3处理次之,且之间无显著差异(P>0.05),NaHSO3处理下靛蓝含量最低,相比于Na2S处理下降了74.52%。而靛玉红含量以Na2SO4和Na2SO3处理次之,且两者无显著差异(P>0.05)。Na2SO4和Na2SO3处理相比于Na2S处理含量下降了26.28%和32.30%。CK、NaHSO3和Na2S2O3处理下菘蓝叶片靛玉红含量较低。说明S2-可以显著促进菘蓝叶片中靛蓝和靛玉红含量的累积。
3 讨论
3.1 硫素形态对苗期菘蓝生长的影响
硫作为植物生长的矿质元素,是继氮、磷、钾后的第四大矿质元素,是植物体内酶化反应活性中心的必需元素,也是植物体内叶绿素、氨基酸、蛋白质、辅酶等的组成成分[2]。在本研究中,以Na2SO4处理后菘蓝株高、叶片鲜干重最高,说明SO42-形式的硫素对于菘蓝植物的生长有显著的促进作用。 这可能与SO42-是植物体内最主要的硫素吸收同化利用形式有关[2]。且在无硫条件下,菘蓝植株生物量值仅低于Na2SO4和NaHSO3处理,并非处于最低水平。这与缺硫或低硫会在一定程度上抑制植物干物质积累不一致[14]。推测原因可能是由于菘蓝除通过根部吸收硫素外,还可通过叶片吸收空气中SO2或H2S气体来补充生长所需的硫素。此外,不同硫素形态下营养液的pH值存在差异,其中CK和Na2SO4处理的营养液呈中性,而NaHSO3处理下偏弱酸性,其余处理(Na2SO3、Na2S和Na2S2O3)呈弱碱性。本研究中硫素形态对菘蓝生物量的影响排序为:Na2SO4>NaHSO3>CK>Na2SO3>Na2S2O3>Na2S。初步推断,以中性或弱酸性环境较适宜于菘蓝的生长,而碱性环境可能会对菘蓝生长有一定的抑制作用。在实际栽培生产中,植物对硫素的吸收利用往往还受较多因素的影响,还有待进一步研究。
3.2 硫素形态对苗期菘蓝光合生理的影响
含硫化合物硫脂是植物光合作用过程中重要组分,硫脂是叶绿体内一个固定的边界膜。硫还是铁氧还原蛋白的重要组分,在光合作用及氧化物的还原中起电子转移作用[15]。因此,硫素对植物光合效率具有重要的作用。在本研究中,无硫处理下(CK)菘蓝叶片中叶绿素含量最低,这可能与缺硫会导致植物叶绿体结构发育不良,使正常的叶绿素合成代谢受到干扰相关。且菘蓝在CK处理下净光合速率同样也处于相对较低的水平,这一方面与CK处理下叶片中叶绿素含量较低有关;另一方面也与缺硫导致菘蓝气孔导度变小有关,使胞间二氧化碳浓度升高,从而限制了菘蓝的净光合速率[16]。此外,本研究还发现硫素处理下,菘蓝的叶绿素含量显著高于对照组,净光合速率除Na2S2O3处理外也均高于CK处理。其中以Na2SO4与NaHSO3处理下菘蓝的光合效率较高,这可能是由于不同形态的硫素在合成有机硫过程中存在一定的差异,而叶片中有机硫主要集中在叶肉细胞的叶绿体蛋白上[17],从而使得不同硫素条件下菘蓝的光合色素累积量与光合作用效率存在差异。
3.3 硫素形态对苗期菘蓝氮、硫代谢的影响
在整个植物体硫同化过程中,不同的硫素存在于各自特定的阶段。因此,不同硫素形态对植物体内硫代谢酶活性有较大的影响。本研究中,Na2SO4处理下菘蓝ATPS和APR活性较高,约为其他处理的两倍。说明SO42-形态的硫素会经过活化氧化阶段形成SO32-,从而促使ATPS及APR活性的提高。但无硫处理下(CK)的ATPS活性显著高于硫素处理下(除Na2SO4处理外),这与徐瑶等[8]发现施硫会提高白菜叶片中ATPS-2、ATPS-3、ATPS-4 表达量结论相反,这可能是因为不同植物对于硫素的吸收同化利用存在区别,且受包括遗传因素和环境条件等共同影响[18]。植物半胱氨酸合成最后一步由2个酶共同催化完成,先由丝氨酸乙酰转移酶(serine acetyltransferase, SAT)催化乙酰辅酶A和丝氨酸生成O-乙酰丝氨酸(O-Acetylserine, OAS),然后H2S和OAS在OAS-TL的催化下形成半胱氨酸。不同形态硫素对于半胱氨酸合成酶的影响大小:Na2SO4>NaHSO3>CK>Na2S>Na2SO3>Na2S2O3。这可能是由于在菘蓝体内,以SO42-形式的硫素同化途径是主要的硫素利用方式,而其他形式的硫会对硫素同化过程形成一个反馈调控,且OAS-TL酶活性还受植物体内OAS含量的影响,而OAS积累到一定程度时,会形成一个可逆的反应系统[19]。因此OAS-TL对于不同硫素形态的响应受多因素的影响。
植物中硫酸盐同化和氮同化是互相协调的,一种元素的缺乏会抑制另一个元素的同化途径。缺硫可能会导致N和C同化的抑制和氨基酸的积累[20]。朱英华等[21]研究发现硫素可以提高烟草(Nicotianatabacum)叶片中硝酸还原酶活性。朱云集等[22]研究发现高氮水平下施硫降低了小麦(Triticumaestivum)叶片中硝酸还原酶的活性,但提高了谷氨酰胺合成酶的活性。而Saccomani等[23]却发现硫素对于玉米(Zeamays)中GS活性的影响不显著。由此可见,氮代谢相关酶活性的强弱不仅受植物生长环境中营养状况的影响,也可能与植物种类、光照、温度、水分等多种环境因素有关[24]。无硫条件下菘蓝叶片中谷氨酰胺合成酶活性最高,而硝酸还原酶活性是CK处理下较高。这可能与缺硫或者低硫条件促进了氮代谢对同化能的竞争有关[22,25],硫素的缺失可以在一定程度上提高氮同化的竞争力从而使硝酸盐和铵的同化增强。而不同形态硫素对于菘蓝的谷氨酸脱氢酶(GDH)活性的影响,以菘蓝在碱性环境下(Na2S2O3,Na2S和Na2SO3)显著高于中性或弱酸性环境(CK,NaHSO3和Na2SO4)。GDH具有双重功能,既能催化NH4+与α-酮戊二酸合成谷氨酸,又能催化谷氨酸氧化脱NH4+[26-27],推测可能是由于酸碱性环境影响植物体内的NH4+含量变化,从而改变GDH酶活性,其具体机制还有待进一步研究。
3.4 硫素形态对苗期菘蓝营养物质及活性成分积累的影响
硫是组成蛋白质的半胱氨酸、胱氨酸和蛋氨酸等含硫氨基酸的重要组成成分,也是多种蛋白质和酶的组分[28]。因此,硫素对菘蓝体内蛋白、氨基酸等营养物质含量有明显的影响。无硫条件下(CK)菘蓝叶中可溶性蛋白和游离氨基酸含量均处于最低水平,说明缺硫会抑制其叶中氨基酸和蛋白质的合成。菘蓝叶中蛋白和氨基酸含量对硫素形态的响应基本呈现游离氨基酸含量高低与可溶性蛋白为相反趋势(除Na2SO3处理外),说明不同硫素处理后,氨基酸与蛋白质之间的合成与分解效率存在差异。而可溶性糖含量变化以S2-和S2O32-处理下含量较低,说明该两者形态的硫素对于可溶性糖的合成可能存在一定的抑制作用。
硫作为植物营养元素之一,除对植物初生代谢产生影响外,对植物次生代谢也具有重要的作用。油菜(Brassicanapus)在缺乏硫素的情况下,芥子油苷含量显著下降,一方面是因为合成减少,另一方面为达到植物自身对硫的需求而促进其分解[29]。靛蓝、靛玉红含量在无硫条件下均未最低,说明在缺硫情况下,菘蓝也可能通过其他途径来平衡营养元素的缺失或通过刺激次生代谢来抵御外界不良环境。此外,以S2-形式的硫素能显著提高靛蓝、靛玉红含量。该现象不同于硫素形态对其生理、生长等的影响,原因可能是在植物营养充足的情况下,植物资源分配存在差异,适宜的硫素形态使营养较多地用于植株生长,而在Na2S处理下,植物硫素资源更多转向了次生代谢,这也是菘蓝对不利环境的一种适应机制。在实际生产中,若是以获取高含量提取靛蓝、靛玉红的药材为栽培目的,可参考施以S2-形态的硫素。
4 结论
在本试验条件下, Na2SO4处理后菘蓝生长状况及生物量积累情况最佳,缺硫对于菘蓝生长抑制作用不显著,CK处理下菘蓝生物量高于其他硫素处理(除Na2SO4和NaHSO3外)。缺硫显著降低了菘蓝叶片中光合色素及光合效率,各硫素处理中,以Na2S处理下菘蓝叶片中叶绿素含量最高,NaHSO3处理的光合效率最高。硫素的缺失及不同硫素形态对于菘蓝叶片氮、硫代谢酶活性影响无明显规律,其中以Na2SO3处理下NR活性最高,CK处理下GS活性最高,Na2S2O3处理下GDH最高,Na2SO4处理下GOGAT最高。对于硫代谢酶(ATPS,APR和OAS-TL)活性以Na2SO4处理下最高。缺硫能抑制可溶性蛋白和游离氨基酸的合成。其中可溶性糖和可溶性蛋白含量以NaHSO3处理下最高,而游离氨基酸以Na2SO4处理下最大。不同于初生代谢,菘蓝叶片中靛蓝和靛玉红含量均以Na2S处理最高。综合不同硫素形态对于菘蓝栽培品质分析,以Na2SO4处理即SO42-形态的硫素最有利于菘蓝的栽培。
