阈值温度和积温对川西高原林线岷江冷杉径向生长的影响
2019-03-22郭滨德王晓春张远东
郭滨德,王晓春,*,张远东
1 东北林业大学林学院生态研究中心, 哈尔滨 150040 2 国家林业局大兴安岭林业调查规划设计院,加格达奇 165000 3 中国林业科学研究院森林生态环境与保护研究所, 国家林业局森林生态环境重点实验室, 北京 100091
林线树木生长对气候响应的敏感性问题比较复杂。在青海阿尼玛卿山,7月低温和多雨是高山林线树木生长的主要限制因子,同时7月温度对林线分布也有影响[9]。在中国东北,温度(尤其积温)是林线树木生长的主要限制因子,降水对林线树木生长的影响不如温度强烈[10]。但也有研究认为,1月平均温度和积温是限制林线树木生存的主要气候因子,温度升高会导致林线上升[11]。在贺兰山地区,干旱是限制东坡高山林线青海云杉(Piceacrassifolia)径向生长的主要因子[12]。也有研究认为在高山林线内小生境的差异性较大,这可能降低树木对温度变化的敏感性[13]。上述研究主要集中在青藏高原地区、长白山地区及贺兰山等地区,然而在川西高原高海拔地区针对岷江冷杉(Abiesfaxoniana)开展树木径向生长与气候因子关系研究则相对较少,尤其缺乏林线附近极端条件下树木生长与气候关系的研究。
岷江冷杉是青藏高原东缘形成林线的主要树种之一,岷江冷杉在海拔3000 m以上区域常以纯林形式存在,是岷江上游地区阴坡林线的主要建群树种[14- 15]。林线岷江冷杉径向生长对气候变化十分敏感,是研究高山树木生长对气候变化响应的理想树种。目前,国内有关高山林线对气候响应研究主要侧重于林线内部植被格局变化、林线位置波动和种群密度变化等方面[5,16-24],而运用树木年轮对高山林线的研究则主要集中在生长与气候关系上[9,13,25- 27]。为了进一步挖掘树木年轮中的气候信息,依据温度连续变化过程,探索温度对林线岷江冷杉径向生长的影响,找出影响林线岷江冷杉径向生长的敏感温度,解释温度累积变化对树木生长的影响非常关键。本研究利用松潘气象站日值温度资料与弓杠岭林线岷江冷杉树轮资料,分析不同温度的起始时间和持续时间的周年变化与年轮特征的关系,尝试从多年日均温度变化中提取对林线岷江冷杉径向生长影响较大的阈值温度(阈值温度主要指生长开始、结束日期的平均温度以及生长季的积温),这对于深入了解影响川西高原高山林线树木生长的关键温度及预测未来全球变暖对高山林线动态变化的影响具有重要参考价值。
1 材料与方法
1.1 研究区域概况

图1 松潘气象站月平均温度和总降水变化(1951—2013年) Fig.1 Variations of monthly mean temperature and total precipitation in Songpan meteorological station (1951—2013)
本研究采样点位于青藏高原东南缘岷山山脉东段弓杠岭林线(33°3′N,103°47′E),海拔3620 m。该区域受西风南支急流、东南季风以及西南季风等综合影响,形成了冬寒夏凉、降水适中的山地气候[28]。年平均气温5.9℃,其中1月份平均气温最低(-3.9℃),7月份最高(14.7℃)(图1),≥10℃的年积温1419.8℃,无绝对无霜期;年平均降水量718 mm,雨季(5月中旬至10月上旬)占全年降水量的82%[28]。研究地点在青藏高原东缘林线亚高山地段,紫外辐射强,气温变化快,昼夜温差大。林线植被是以岷江冷杉为主的高山针叶林。土壤主要是山地暗棕壤,土层较厚,地表凋落物较少[28, 29]。
1.2 样品采集
2014年7月在川西高原九寨沟县弓杠岭林线附近,选择生长良好、年龄较老且受人为影响较小的区域进行岷江冷杉树木年轮的取样工作。取样时用内径为5.15 mm的生长锥在胸高处(1.3 m)钻取树芯,尽量取到髓心。每株树采集2个年轮样芯,最终共获得40根样芯[28]。将取到的样芯装入塑料管内,并进行编号(表1)。

表1 林线岷江冷杉树轮年表采样点信息
1.3 数据处理
将年轮样芯带回实验室后,按照Stokes和Smiley[30]阐述的方法进行预处理,经晾干、固定、打磨后,在双筒显微镜下用骨架图法进行目视交叉定年。然后用Velmex年轮测量仪测量年轮宽度,精度可达到0.001 mm。测量后的树轮样本序列用COFECHA程序对定年和测量结果进行检验[31],并消除定年和宽度测量过程中出现的误差,确保年轮的日历年代准确。经过交叉定年的年轮序列,利用ARSTAN程序[32]中的负指数或线性函数进行去趋势和标准化,目的是消除树木生长过程中与年龄增长相关联的生长趋势,以保留更多的气候信号。采用双权重平均法对年轮曲线进行标准化,并最终建立弓杠岭林线岷江冷杉年表。从年表统计特征可知,标准年表具有较高的质量,因此本研究利用标准年表进行相关分析[28]。
1.4 气象资料与统计分析
由于九寨沟县缺乏长期气候数据,本文根据采样点的位置和海拔高度,选择距离取样点较近的松潘气象站(32°39′N,103°34′E,海拔2851 m)1951—2013年的气候数据用于分析。研究中所用气候资料为日平均气温、月平均气温和总降水量。生长季前(Before growing season, BG)定义为当年的3—4月,当前生长季(Current growing season, CG)定义为当年5—9月。由于树木生长不仅与当年的气候因子有关,还与上一年气候因子有关,因此选择上一年9月到当年9月共13个月的气温和降水量与岷江冷杉年表进行相关分析,同时分析岷江冷杉与生长期其他参数(初日、终日、持续时间和活动积温)的相关性。年表与气候要素之间的相关分析用SPSS 19.0软件计算,作图用SigmaPlot 12.5 软件完成。
1.5 日值温度资料处理
采用5日滑动平均法确定1951—2013年稳定通过某一温度界限值的初日和终日,并确定初日和终日在一年中的位序,计算稳定通过某一温度的活动积温和持续天数[33]。树木径向生长光合作用的下限温度为5℃,因此计算活动积温的限制温度从5℃开始,每隔0.5℃计算1次,一直到15℃。这样就得到生长期活动积温、持续天数、初日和终日序列[28]。
为了较准确地确定5℃至15℃温度范围内对林线岷江冷杉生长更为重要的阈值温度,我们对5—15℃不同阈值温度初日、终日日期与岷江冷杉径向生长的相关系数进行了Mann-Kendall检验,通过突变点和趋势分析来判断更为重要的阈值温度。Mann-Kendall检验用Matlab编写的小程序完成(需要者可从通讯作者处获得)。
2 结果与分析
2.1 林线岷江冷杉年表统计特征
林线岷江冷杉年表长度为204年;一阶自相关系数相对较高,说明前一年的气候对当年的树木生长也有影响,高海拔林线岷江冷杉年轮序列中含有较高的低频变化信息[34];样本代表性较高(表2),说明所采样本可以代表采样点树木变化的总体特征。以上年表统计特征表明该研究地区的岷江冷杉树轮资料适合用于树轮气候学研究。
在 《西行漫记》中,斯诺谈到中国共产主义运动和共产国际,他明确指出共产国际在中国革命中的作用被大大夸大了。中国革命的产生和发展无疑受到俄国和共产国际的影响和帮扶,但共产国际的作用并非决定性的。 “有一段时期,中国共产党同共产国际失去了联系,共产党人在自己的领导下自发组织苏维埃运动。而相比于美国对南京政府的援助,共产国际给予中共的实际财政援助实在是少得惊人。”[1]343-344在这种力量对比悬殊的条件下,中国革命能取得胜利,靠的正是中国共产党从中国实际出发,坚持独立自主地领导中国革命。

表2 林线岷江冷杉标准年表的主要统计特征
Note:SSS-子样本信号强度, subsample signal strength

图2 林线岷江冷杉标准年表的变化趋势 Fig.2 Variations of the tree-ring standard chronologies of treeline Abies faxoniana
由图2可以看出,林线岷江冷杉年表的整体变化趋势在1980年前后发生明显改变,1980年前林线岷江冷杉生长保持稳定,而1980年后林线岷江冷杉生长明显加速(0.03a-1,r2=0.71)。
2.2 林线岷江冷杉生长-气候关系
由林线岷江冷杉生长-气候关系可以看出(图3),温度是林线岷江冷杉的主要限制因子。其中林线岷江冷杉径向生长与生长季(5—9月)、冬季(12、1和2月)及上一年生长季末期(9、10月)温度显著正相关(P< 0.05)。林线岷江冷杉与降水的相关性明显弱于温度,仅表现在与6月降水显著负相关(P<0.05)。
2.3 林线岷江冷杉与生长期其他参数的相关分析
林线岷江冷杉径向生长与不同起始温度的初日时间呈负相关,说明在5—15℃的范围内,初日时间越早对林线岷江冷杉径向生长的促进作用越大。其中,9.5℃的初日时间对林线岷江冷杉的负相关性最强(P<0.05);林线岷江冷杉径向生长与不同起始温度的终日时间则表现为正相关,说明终日时间延迟会对林线岷江冷杉有促进作用(图4)。其中,6.5℃的终日时间与林线岷江冷杉的正相关性最强(P<0.05)。

图3 林线岷江冷杉年轮指数与气候因子的相关分析 (虚线表示95%置信水平)Fig.3 Correlation coefficients between tree-ring standard chronologies of treeline Abies faxoniana and monthly climatic factorsBG: 生长季前(3—4月) Before growing season (March—April); CG: 当年生长季 (5—9月) Current growing season (May—September). 虚线表示95%置信水平,负号代表前一年 Dotted line represented 95% confidence level, and minus before the figures in the x-axis represented the previous year

图4 林线岷江冷杉标准年表与生长期初日和终日的相关性Fig.4 Correlation coefficients between tree-ring standard chronologies of treeline Abies faxoniana and the first and last dates of the growing season at various temperature thresholds
活动积温表示一定温度期限内的热量积累,是表示某地热量条件的指标[35]。由图5可以发现,在5—15℃范围内,林线岷江冷杉径向生长与不同起始温度的生长期活动积温和持续时间呈正相关,其中林线岷江冷杉径向生长与6.5—9.5℃之间的生长期活动积温及9.5℃持续时间正相关性最高。说明林线岷江冷杉径向生长的下限温度在6.5—9.5℃左右 (图4、图5)。
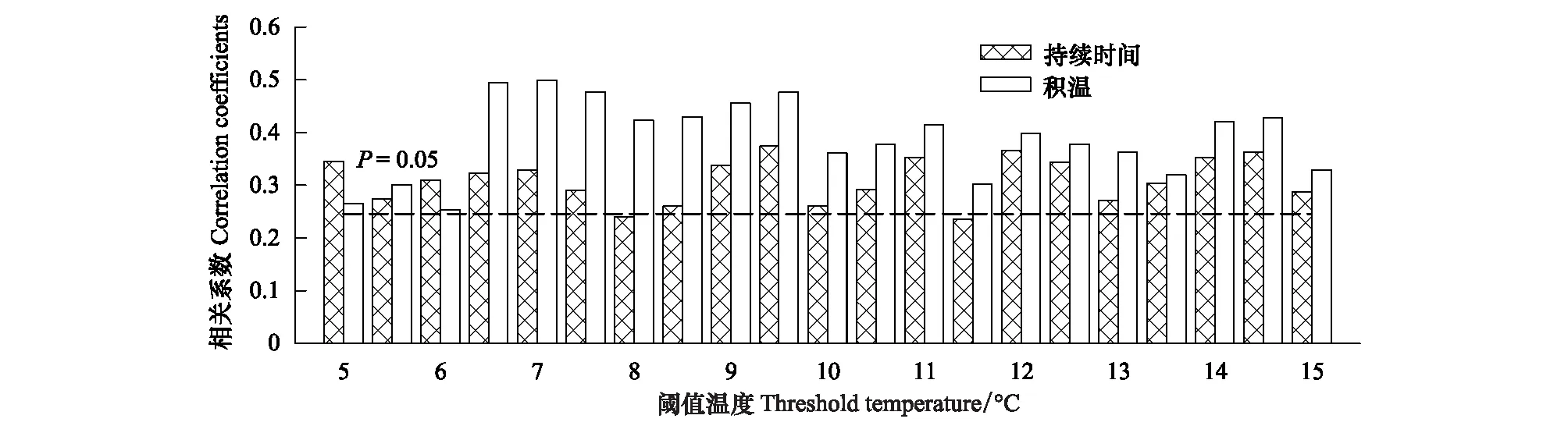
图5 林线岷江冷杉标准年表与生长期活动积温的相关性Fig.5 Correlation coefficients between tree-ring standard chronologies of treeline Abies faxoniana and the cumulative temperature and continuous days of growth season at various temperature thresholds
生长季初日、终日日期与林线岷江冷杉径向生长相关系数的Mann-Kendall检验表明,生长季初日的阈值温度突变点在7—8.5℃(图6);而生长季终日在11.5℃是显著的突变点,在9.5℃时几乎显著相交 (图6)。Mann-Kendall检验可能表明林线岷江冷杉在生长季初期开始活动的阈值温度和生长季末期停止生长的阈值温度可能存在差异,但这是否仅是突变检验的结果,还是生理上就存在这样的差异,还需要生理实验进行验证。结合图4、图5的相关分析认为,弓杠岭林线岷江冷杉生长的初日温度在7℃,而生长停止的阈值温度在9.5℃ (图6)。
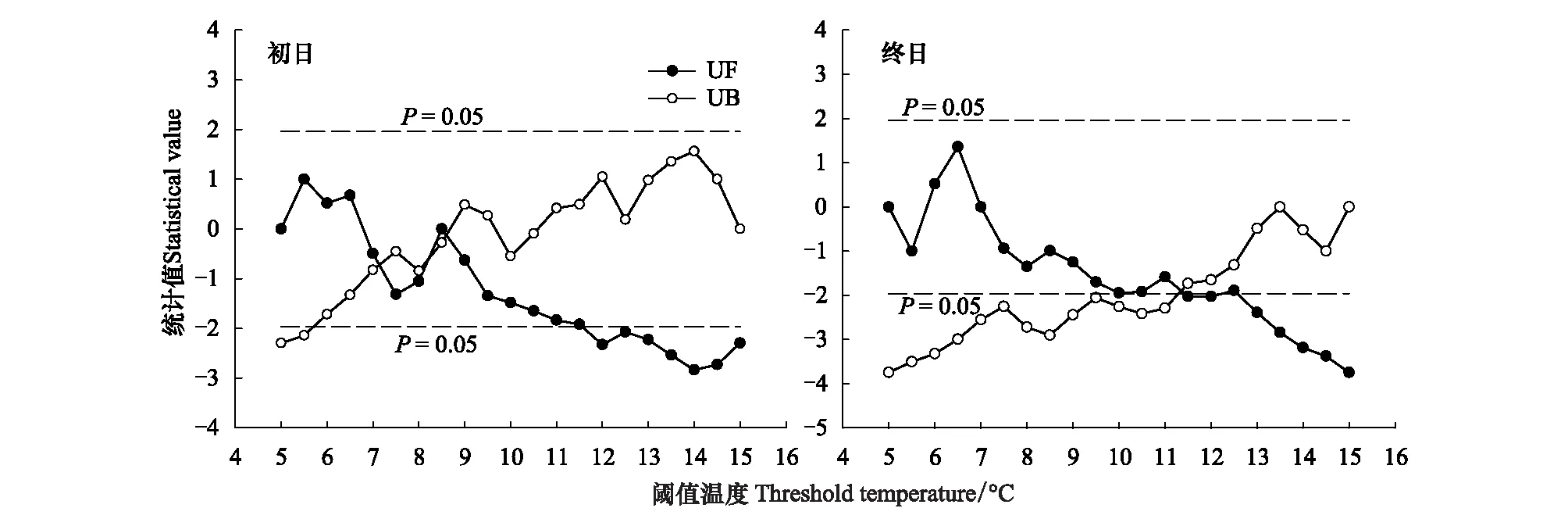
图6 林线岷江冷杉生长期初日(A)、终日(B)日期与年轮指数相关系数的MK检验Fig.6 Mann-Kendall test of the correlation coefficients between the tree-ring index of Abies faxoniana at treeline and the initial (A) and final (B) date of the growing season at various threshold temperatureUF和UB分别代表初日、终日日期与年指数相关系数进行MK检验的正向和逆向变化曲线
弓杠岭地区多年日平均气温从生长季开始时的7℃到生长季结束时的9.5℃,持续天数从第108天到第274天,表明弓杠岭林线岷江冷杉生长期为4月中下旬到10月初(图7),在林线岷江冷杉年表与月气候因子相关分析中也发现,5月到9月温度对林线岷江冷杉径向生长有显著的促进作用。

图7 松潘气象站多年日平均气温变化 Fig.7 Variation of the daily mean temperature at Songpan (1951—2013)
由图8可以看出,1980年前生长季温度和积温变化趋势不明显,但1980年后生长季温度以0.42℃/10a的速率快速升高,而积温以118.8℃/10a的速率快速升高,随着温度升高(1980年后)林线岷江冷杉径向生长显著增加(图2)。林线岷江冷杉生长期初日显著提前,每10年变化幅度为4.6d/10a (r2=0.19,P=0.01),生长期终日年际变化趋势较弱,每10年变化幅度为1.8d/10a (r2=0.04,P=0.25),生长期持续天数整体上呈显著上升趋势,每10年变化幅度为6.4d/10a (r2=0.19,P=0.01) (图8)。

图8 1980年升温前后林线岷江冷杉生长期气候参数变化比较Fig.8 Variation of the climatic parameters during the growing season at Songpan before and after 1080
3 讨论
3.1 林线岷江冷杉对月气候条件的响应
在林区,一般随海拔升高降水增加,但温度则明显下降。在川西弓杠岭林线区域,温度是林线岷江冷杉径向生长的主要限制因子,这与Gou等[36]在祁连山研究结果一致。弓杠岭林线岷江冷杉径向生长与生长季温度显著正相关,这与川西高原地区[37- 38]、青藏高原地区[39]及长白山地区[40- 41]等的研究结果相类似。树木径向生长与温度关系是树木生理特征与环境条件长期适应的结果[42]。森林上限温度较低,生长季相对较短,树木年轮一般会在生长季7—8月热量条件较好的50天内形成[4]。5—9月生长季是林线岷江冷杉生长最旺盛的时期,此时较高的温度有利于增加光合作用,提供充足的光合产物供树木生长,加之林线附近生长季昼夜温差大,降低了呼吸消耗,从而使得营养物质净积累增加,促进林线岷江冷杉径向生长[34]。林线岷江冷杉与生长季前(3—4月)温度显著正相关,这主要是由于生长季前温度偏高有利于加速积雪融化,为形成层细胞分裂提供充足的营养物质,同时较高的温度可以相应地延长植物的生长期,促进树木生长[43- 45]。在高海拔地区,温暖的冬季可以避免叶组织冻结[34],保证树木生长正常的生理代谢活动,使得树木在下一个生长季的径向生长加快[44],因而冬季温度对林线岷江冷杉有显著的促进作用,这与其他高山林线[46- 48]的研究结果相似。林线岷江冷杉径向生长还受上一个生长季末的温度影响,这可能是上一年生长季末温度适宜有利于光合产物积累,进而给下一年冷杉径向生长创造了良好的物质条件[49-51]。
3.2 林线岷江冷杉径向生长的阈值温度
年轮是形成层周期性分裂形成木质部的结果[52],木质部的形成具有一定的节律性[53],春季形成层开始活动的时间主要受到温度的限制[54],只有当春季外界温度或某一温度的积温超过一定阈值时形成层才结束休眠开始活动[55- 56],即形成层活动存在温度阈值。在欧洲和加拿大的寒冷气候区,尽管采样地点和树种不同,但针叶树形成层开始活动的平均温度在8—9℃[57]。而中国北亚热带地区,10.5℃是马尾松木质部形成层活动的敏感温度[58]。与上述研究结果类似,林线岷江冷杉径向生长与9.5℃活动积温及生长期持续时间显著正相关(图5),这可能说明9.5℃是林线岷江冷杉形成层活动的敏感温度,春季温度高于7℃之后光合作用逐渐增强,形成层活动开始加速,形成层细胞不断分裂使形成层区细胞数量增加,当秋季温度低于9.5℃之后形成层活动停止,此后已分化的细胞继续进行细胞伸展、次生细胞壁加厚和木质化,来完成细胞的生长[59]。多年日平均温度春季高于7℃至秋季低于9.5℃的天数是第108天到第274天,从形成层开始恢复活动到最后一个新形成的细胞完成生长这个阶段被称为植物的生长期[59],由此可以推断林线岷江冷杉的生长期是4月中下旬至9月末。林线岷江冷杉径向生长与9.5℃的初日时间显著负相关,而与9.5℃的终日时间相关不明显(图4)。这主要是由于温度可以影响生长季前期形成层的活动,改变形成层活动的时间、细胞的分裂速率及活动周期,进而对木质部各阶段细胞分化的开始和持续时间产生影响,但是生长季后期木质部的形成(如次生细胞壁的加厚和木质化的完成)受温度的影响并不明显[59, 60]。林线岷江冷杉径向生长与5—7℃的终日时间显著正相关,这说明5—7℃可能是光合作用的敏感温度,这与袁玉江等[61]、封晓辉等[58]的研究结果相似。一般认为5℃是植物光合作用的最低温度,但光合作用的最低温度会随着不同的树种和地区而发生变化[58]。岷江冷杉是高海拔常绿树种,春季温度超过一定温度后树液开始流动,随着温度进一步升高,光合速率增加,光合产物积累增多[62],林线岷江冷杉进入径向生长旺盛期,9月末林线岷江冷杉形成层活动减缓,生长变慢,当温度低于7 ℃时,形成层活动逐渐停止,树木停止生长[28]。
3.3 林线岷江冷杉径向生长的积温
工业革命以来,全球升温已经成为不争的事实,川西高原大部分地区与全球温度变化相一致,表现出温度显著升高趋势[63]。温度升高会提前和延后树木生长的开始和结束时间[64]。20世纪中期显著升温以后,全球范围内多数植物出现春季物候期提前,秋季物候延后现象[65- 69],本研究也有类似发现。川西岷江冷杉生长季天数显著增加 (6.4 d/10a,r2=0.19,P=0.01),这可能是由于早春温度升高使冷杉春季物候提前导致的[64]。植物不但需要在高于一定的温度后才能生长发育,而且需要累积一定的温度才可以完成其整个生长发育过程,岷江冷杉生长期长度与活动积温存在显著的相关性[70]。1978年后我国大部分地区生长期活动积温和持续天数均出现增加,初日和终日出现提前或者延后现象[71]。研究表明,随着温度升高,生长季延长会对林线岷江冷杉径向生长产生促进作用,这可能会使年轮宽度增加,幼苗更新增多,进而林线上移。
