刺槐人工林无性更新幼苗空间格局及生长特征研究
2019-03-21
(1) 山东省林木种苗和花卉站,250014,济南; 2) 山东省林业监测规划院, 250014,济南;)
森林无性更新是利用树木地下茎和残留体(树桩)上的休眠芽或不定芽形成新的植株的过程,以达到森林更新的目的[1],无性更新是森林更新的重要方式.由于萌生植株能通过其原有的强大根系,更有效地利用土壤中的养分资源,通常比实生植株生长快,具有更强的环境适应能力;能较快地再次占据林窗,在种群竞争中处于有利地位.
刺槐(RboniapsuedoaaccaiL.)属于喜光树种,在温暖湿润的气候条件下能发展人工林,我国刺槐人工林面积超过100万hm2.山东省刺槐林面积17.28万hm2,木材蓄积量375.12万m3,分别占全省阔叶林面积的58%和蓄积量的50%以上,是山东省阔叶树中造林面积最大、木材蓄积量最多的树种之一.刺槐实生林数量成熟龄一般为30-50年,由于其萌芽、萌蘖能力强,生产中一般采取伐后萌芽、萌蘖更新,即在采伐后留桩萌芽更新、或伐桩剥皮(或挖桩)根系萌蘖更新,并于更新后1~3年内通过抚育确定适宜的林分密度[2,3].但无性更新刺槐林速生期维持年限短,后期生长迟缓,干形弯曲,难以成材.及时了解并掌握刺槐人工林萌芽(萌蘖)更新和生长特性,对制订刺槐人工林经营管理方案具有重要意义.当前,对刺槐人工林更新研究主要集中更新复壮措施[4]、萌生苗早期生长规律[5]、更新方式等方面[6-8].但关于刺槐萌生林的幼苗更新格局[9]、萌芽和萌蘖更新幼苗生长[10]缺乏深入了解.本文通过对现有无性更新刺槐林幼苗的调查,比较刺槐人工林萌芽、萌蘖更新幼苗更新空间分布格局及生长特征,以期为刺槐林人工林长期经营提供理论依据.
1 研究区概况
沂山位于鲁中沂蒙山区的北部,东径118°36′~40′,北纬36°10′~13′.沂山在清朝时,曾为放马场,原始植被早已破坏殆尽.沂山林场于1948年建立,通过大规模营建人工林形成以刺槐纯林和松(油松、赤松、黑松)-刺槐混交林为主的植被类型.该林场地处温带季风气候区,年平均温度10.8 ℃,年降水量845 mm.以花岗片麻岩为主,砾岩较少,风化好,母砾疏松,土质类型为山地棕壤,土壤厚度80 cm以上.
2 研究方法
2.1样方调查在刺槐皆伐迹地内置大小为4×20 m标准样地,采用相邻网格线法把调查样地划分成20个2×2 m的小样方.对标准地内各样方的刺槐人工林无性更新的萌芽、萌蘖幼苗和伐桩进行每木检尺和定位,测量出幼苗的地径、高度、坐标以及伐桩的具体位置.
2.2数据处理种群的分布格局是由种群本身生物学特性、生境条件以及两者之间的相互作用所决定的,是对种群个体水平空间上配置和分布状态做出的定量描述,大致分为均匀型、随机型、聚集型3种.刺槐人工林无性更新幼苗内分布型格局运用的检验指标是方差/平均数比率,即为S2/m,记为C,在选择计算方法时,方差均值比率法是建立在严密的数学检验基础上的,其结论不存在难以判断的临界点,分析结果具有相对客观性,比较可靠,是研究植物种群分布格局常用的方法[11].其中,
m=∑f(x)/n;S2={∑(f(x))2- (∑f(x))2/n}/(n-1)
x:样方中某个体数,f:含x个体样方出现的频率,n:样方总数
若:C>1,属于聚集分布,C=1,属于均匀分布,C<1,属于随机分布.
用Excel分析工具库中的双样本方差分析、双样本等方差检验或双样本异方差进行F检验和两样本均值t检验.
3 结果与分析
3.1刺槐无性更新幼苗空间分布格局根据幼苗和伐桩的坐标,可得到刺槐萌芽、萌蘖幼苗以及伐桩的相关分布格局(图1).从图中可见,刺槐由无性更新方式产生的幼苗依伐桩而呈现聚集分布,而萌蘖更新幼苗因主要在刺槐发达的根系上均匀分布.根据各萌芽、萌蘖个体数样方出现的频率计算方差/均值的比值(表1),C(萌芽)=3.013>1符合聚集分布的类型;C(萌蘖)=1.073≈1属于均匀分布的类型.产生这种分布格局的主要原因在于刺槐人工林皆伐迹地的伐桩的保留抑制萌蘖幼苗的生长,促进萌芽幼苗的生长,从而导致萌芽幼苗在伐桩及伐桩周围成团状分布,只有去伐桩、粗根才能促进萌蘖幼苗的发生;萌蘖幼苗均匀分布主要是由其幼苗种群内的个体竞争所引起的,体现在刺槐伐桩根系上的萌蘖幼苗对根系养分的竞争上.聚集分布是刺槐幼苗更新最为常见的分布型,其形成原因包括环境资源的分布不均匀,刺槐幼苗以母株为扩散中心在皆伐迹地内聚集分布.
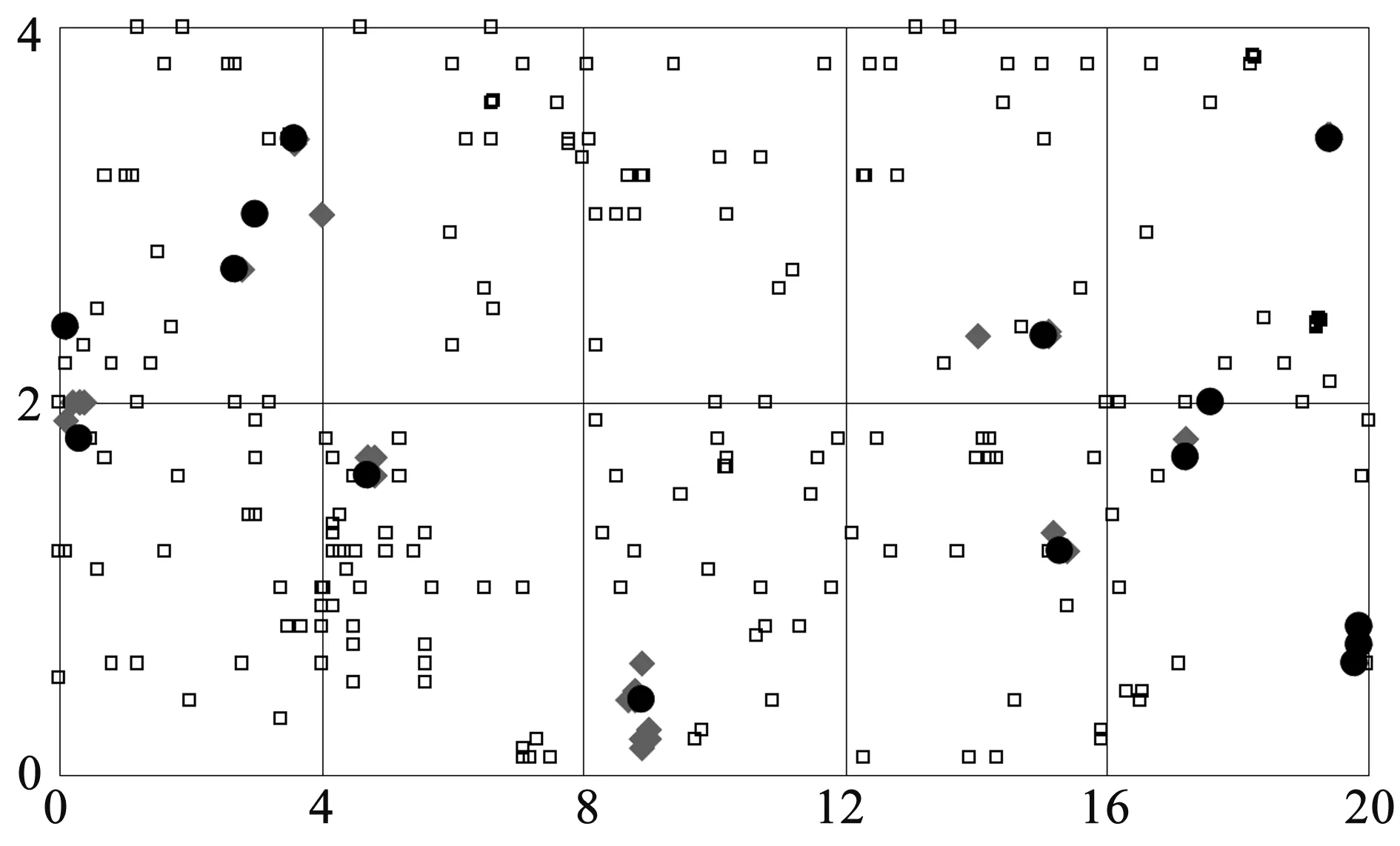
图1 刺槐皆伐无性更新方式及幼苗空间分布格局
3.2刺槐无性更新幼苗地径分布特征
3.2.1 刺槐无性更新幼苗地径分布曲线分析 从幼苗地径的分布情况来看(图2),萌芽幼苗的地径主要分布在0.5~3.0 cm内,占比例88.68%,地径4.5 cm以上的个体极少,且在整个地径分布范围内各区间数量呈现出周期性的波动地:在萌芽幼苗的更新早期,其更新数量年年递增到一个高峰期后骤降,并展开新一轮的动态更新,因为幼苗数量的过多或过少都不利于幼苗的萌生.因此,在刺槐的皆伐迹地上,幼苗的更新过程中受到了不同程度的干扰.这其中包括降水量、个体间竞争、伐桩养分供给等因素的影响导致萌芽幼苗的地径生长速度参差不齐.

表1 调查样方刺槐更新幼苗频率方差均值比率分析

图2 刺槐萌芽和萌蘖幼苗的地径分布
萌蘖幼苗的地径主要集中在0~2.5 cm范围内(图2),占比率95%,地径4.5 cm以上的个体极少.从其数量更新的趋势来看,萌蘖幼苗是逐年递增的.萌蘖幼苗地径分布特征可能受刺槐更新方式的影响,使刺槐萌蘖幼苗直径大多较小,萌蘖幼苗的地径分布图显示出前几年的萌蘖更新由于受到养分的限制,更新频度和速度较为缓慢.近两年,状况得到缓解后,其更新频度有了大幅度的提升.综合萌芽和萌蘖幼苗的地径分布趋势来看,在皆伐迹地更新的初期,由于所需的生境因子的条件不满足导致萌生幼苗的数量极少;萌芽幼苗更新的高峰期比萌蘖幼苗更新的高峰期来得要早;两种更新方式的幼苗地径普遍偏低且分布曲线异于一般的天然更新分布情况.
3.2.2 标准地内无性更新幼苗地径方差检验 在标准地内,由于萌芽和萌蘖幼苗的株数相差较大,需进行双样本方差分析来观测两种更新方式对方差的影响,再进行下一步的均值比较分析,对萌芽和萌蘖地径做t检验(表2).双样本方差分析结果表明在标准地内萌芽和萌蘖地径方差显著,经双样本异方差分析检验两种更新方式的均值有显著的差异且萌芽的地径明显大于萌蘖.说明萌芽幼苗在地茎生长方面有更好的优势.
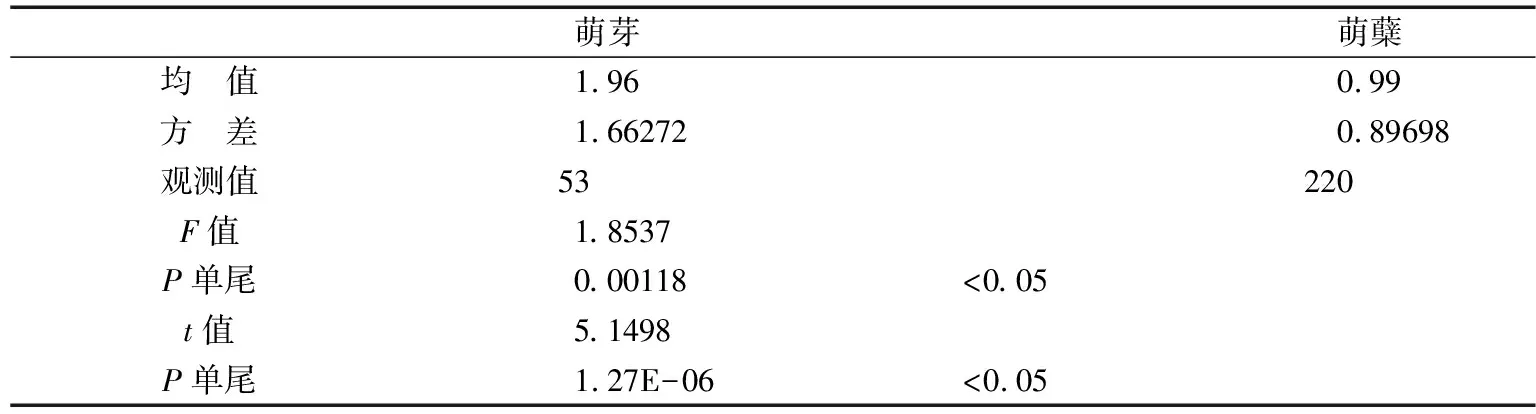
表2 双样本方差分析和双样本方差假设检验(t检验,α=0.05)
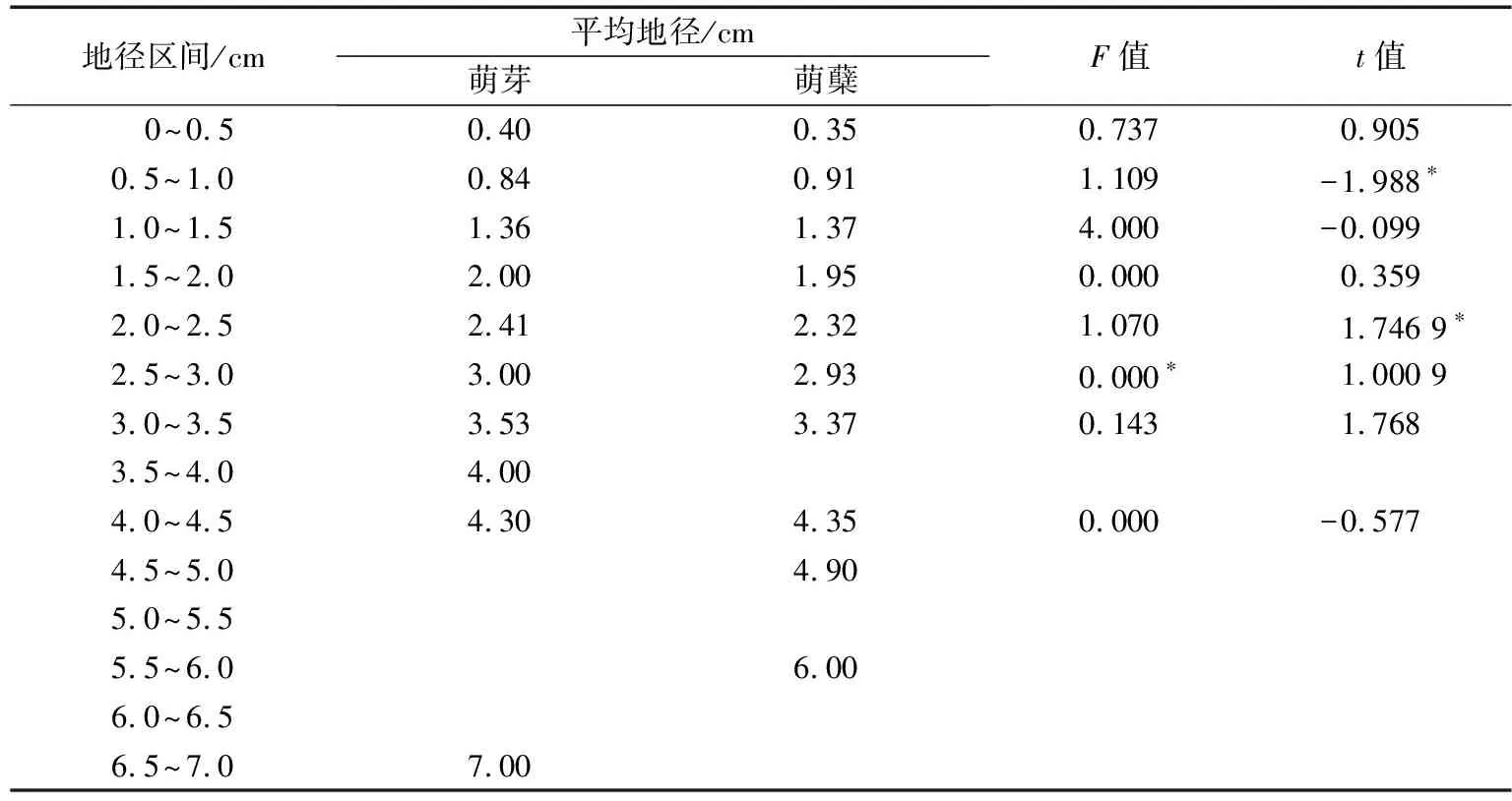
表3 幼苗地径双样本方差分析
注:*代表差异显著,下同.
将标准地内的萌芽和萌蘖有的地径划分为14个分段(表3),3.5~4.0,4.5~5.0,5.5~6.0,6.5~7.0这4个分段的地径值都单一的分布在萌芽、萌蘖、萌芽更新幼苗内,5.0~5.5,6.0~6.5区间则没有更新的幼苗,说明地径值的增大,幼苗的更新方式出现两极化.在两种更新方式都有的区间段:2.5~3.0的分段两方差不齐性,但t值检验结果没有显著差异;0.5~1.0和2.0~2.5区间内均值检验有极小的差异;其他区间的均值检验结果则无明显差异.与总的地径方差分析结果相比较,可以判定出萌芽和萌蘖幼苗地径的差异主要源于一些大龄的幼苗,即在幼苗更新的初阶段,外界因素的干扰或是伐桩养分的供应对于更新方式的影响较为明显.
3.3刺槐无性更新幼苗高度分布特征
3.3.1 刺槐无性更新幼苗高度分布曲线分析 刺槐幼苗不同高度级内的株数分布比例可以作为龄级估测的一个参数.用更新幼苗的高度划分不同等级以代表不同生长阶段分析更新幼苗的动态(图3),Ⅰ级:苗高≤1 m;Ⅱ级: 1 m< 苗高≤2 m;Ⅲ级:2 m<苗高≤3 m;Ⅳ级:3 m< 苗高≤4 m;Ⅴ级:4 m<苗高≤5 m;Ⅵ级:5 m<苗高≤6 m.从图中可以看出萌芽的高度集中在Ⅱ、Ⅲ、Ⅳ区间内,占比率88.67%;萌蘖幼苗则在Ⅰ、Ⅱ区间最为集中,占比例83.64%.若以刺槐幼苗更新的高度生长情况分析,刺槐的萌芽小苗和大苗数量较少,各龄级分布不均.从萌蘖幼苗的分布情况来看,中苗的数量较少,且大苗几乎没有,幼苗集中分布在低龄级.说明刺槐幼苗更新的过程中受到的阻碍较大.近年萌蘖幼苗生长所需的各项因素有所改善,使得其更新数量有了大幅度的提升.中苗和大苗所占的百分比可以看出,早些年皆伐迹地主要以萌芽的方式更新,而受养分竞争和土壤湿度的影响萌蘖幼苗更新十分缓慢.经过双样本方差分析(表4),标准地内萌芽和萌蘖高度的方差不齐性,且双样本异方差的检验结果表明萌芽、萌蘖的高度均值有极显著的差异,萌芽的高度大于萌蘖的高度.

图3 刺槐萌芽和萌蘖幼苗高度级数量分布

萌芽萌蘖均 值2.4091.420方 差1.0720.758观测值53220F值1.4132P单尾0.042<0.05t值6.429P单尾6.42E-09<0.05
3.3.2 标准地内高度分段方差检验 在给高度划分的前5个区间内,方差均为齐性.但在Ⅱ小分段,即萌芽幼苗生长的第二个高峰期、萌蘖幼苗的第一个高峰期,萌芽和萌蘖的更新幼苗高度均值差异显著,且萌芽幼苗稍高于萌蘖幼苗;在Ⅵ区间的高度值只有萌芽幼苗;余下分段的高度均值则无明显的差异.由双样本方差分析结果(表5)可以初步判断出近几年更新方式对幼苗的高度的影响不显著;在各种环境因子均有利的情况下萌芽幼苗仍然保持着生长的优势;在初期更新方式以萌芽为主,即萌芽要比萌蘖幼苗更新的早些,这一点是与地径分布曲线的结果分析是相一致的.

表5 幼苗高度双样本方差分析
4 结 语
刺槐萌芽更新与根蘖更新相比,前者生长快,但密度小、更新频度低.刺槐萌芽幼苗靠伐桩的庞大根系提供养分和水分,使得幼苗时期的萌芽幼苗比萌蘖幼苗生长快.而萌蘖幼苗的生长集中在刺槐伐桩的细根上,伐桩和粗根的保留阻碍了萌蘖幼苗的生长.随着时间的推移,可以发现更新方式对幼苗的高度和地径影响并不明显;但在萌发幼苗的数量上因密度效应、水分和养分的供给不足等问题出现了不同程度的分化.
在调查的标准地内刺槐萌芽幼苗以聚集分布的形式出现;萌蘖幼苗整体呈现均匀分布.这种分布格局的出现可能与刺槐人工林皆伐迹地立地条件有关,因为伐桩的保留一方面促进了刺槐萌芽幼苗的生长但另一方面抑制了萌蘖幼苗生长.两种更新幼树相互嵌套,便于更新林分密度的人为控制,使林分尽快形成较为合理的群体结构持续发挥改善生境、保持水土之功效.若以地径为生长指标判断幼苗更新状况,可发现萌芽幼苗优于萌蘖幼苗且萌芽幼苗的地径和高度的高峰期要比萌蘖幼苗来得早些.地径和高度的阶段性分析结果表明萌芽和萌蘖幼苗的地径和高度的均值差异主要源于萌发苗始期,因为萌芽比萌蘖幼苗提前10~30 d.
资源分配假说认为,萌生更新能力取决于母树地上器官和地下器官的资源储存.地上部分损失后,树桩和根系是树木萌生更新所需营养的主要来源,其中根系的资源储存量对于树木萌生再生具有重要的意义.有研究证实,在植株茎干砍伐后的萌生过程中,植株储存的碳水化合物大量消耗用于萌芽更新幼苗的生长[12].总体来说,若以幼苗的更新质量作为标准,萌芽的更新方式较为优良.但在更新的数量上萌蘖要远远大于萌芽的更新幼苗.故而,应根据材种要求和品质确定合适的无性更新方式.林地和伐桩处理的越彻底,萌生苗越均匀,越能促进萌蘖幼苗的早期生长.刺槐林萌生更新时,应根据林地母树数量、林地海拔、坡度、土层厚度等因子选择处理伐桩的适宜措施.若环境条件较好,从细根上产生的根蘖苗能很快的形成独立根系,后期生长并不次于留桩剥皮处理,生产上有希望培育成林相整齐、形质优良的刺槐萌芽林.
